
Авторы: Thais A. C. Nogueira, Igor Luis Kaefer, Marco A. Sartim, Manuela B. Pucca, Jacqueline Sachett, André L. Barros, Moysés B. A. Júnior, Djane C. Baía-da-Silva, Paulo S. Bernarde, Hector H. F. Koolen и Вуэлтон М. Монтейро.
Received: 18 July 2022; Accepted: 24 August 2022;
Published: 06 October 2022.
Ключевые слова: двуцветная древесная лягушка, гигантская листовая лягушка, гигантская обезьянья лягушка, капум, кампо, двухцветная листовая лягушка, vacina do sapo, лягушачья вакцина
Phyllomedusa bicolor (Phyllomedusidae), известная в Бразилии как камбо, — древесная лягушка, широко распространенная в странах Южной Америки и известная тем, что выделяет кожный секрет, богатый биологически активными пептидами, которые часто используются в ритуалах коренного населения. Биологическое действие кожного секрета было отмечено в первых исследованиях с участием коренного населения. В течение последних шести десятилетий исследователи детально изучают химический состав, а также потенциальное фармакологическое применение входящих в него компонентов. По этой причине коренные общины и представители здравоохранения опасаются неправильного использования камбо, что может привести к осложнениям со здоровьем или даже к смерти пользователей. Цель данной статьи — представить трансдисциплинарный обзор, объединяющий знания о биологии P. bicolor, этнознания о ритуале камбо, химию и фармакологию кожного секрета этого вида, а также медицинские аспекты неизбирательного использования камбо. Кроме того, данный обзор призван пролить свет на перспективы дальнейших исследований, связанных с камбо.
История Phyllomedusa bicolor
1.1 Виды: Общие аспекты
В странах Южной Америки Phyllomedusa bicolor известна как двухцветная древесная лягушка, гигантская обезьянья лягушка, гигантская листовая лягушка, двухцветная листовая лягушка, восковая обезьянья лягушка, камбо (а также кампо) (Frost, 2021). В литературе слово kambô обычно имеет два значения. Оно может использоваться как для обозначения древесной P. bicolor, так и для шаманского ритуала коренного населения (Caramaschi and Cruz, 2002; Frost, 2021). Кожный секрет лягушки на протяжении веков использовался коренными жителями Западной Амазонии в шаманских ритуалах исцеления и очищения, известных как камбо (Erspamer-Falconieri et al., 1970) или капум, или «вакцина от жабы» (vacina do sapo, по-английски). Однако эта практика распространилась на городские центры в качестве альтернативы традиционной медицине (Silva et al., 2019). Это фармакологически важный класс, поскольку он считается богатым источником биологически активных пептидов, многие из которых уже идентифицированы или охарактеризованы, хотя, по оценкам, существует множество соединений, которые еще предстоит открыть (Thompson and Williams, 2022).
Phyllomedusa bicolor была классифицирована Боддаертом из Суринама (1772) как Rana bicolor, а позднее Ваглер (1830) предложил для этого вида род Phyllomedusa. Таким образом, P. bicolor является типовым видом своего рода. Название Phyllomedusa происходит от греческих слов «phyllo» (лист или листва) и «medousa» (королева или защитница), что означает «королева листвы» или «хранительница листвы» (Caramaschi and Cruz, 2002). Специфическое название «bicolor» связано с двумя основными цветами этой лягушки (Boddaert, 1772): спина темно-зеленая, а брюхо от белого до желто-белого или кремового. Монофилия рода Phyllomedusa решительно поддерживается Duellman et al. (2016), и в настоящее время род насчитывает 16 классов (Frost, 2021), причем Phyllomedusa chaparroi является самым недавно описанным (Castroviejo-Fisher et al., 2017). Phyllomedusa bicolor и Phyllomedusa vaillantii часто восстанавливаются как родственные виды, которые объединяет наличие остеодерм в дорсуме кожи (Ruibal and Shoemaker, 1984; Faivovich et al., 2005; Wiens et al., 2006; Moen and Wiens, 2009; Faivovich et al., 2010; Duellman et al., 2016).
1.2 . Сезонность, репродуктивная биология и жизненный цикл
Размножение происходит в основном в сезон дождей, а продолжительность размножения варьирует в географических пределах распространения вида, а также из-за участившихся экстремальных климатических явлений, таких как Эль-Ниньо и Ла-Нинья (Lima et al., 2012; Rocha et al., 2021). В Центральной Амазонии размножение происходит в основном с ноября по май (Lima et al., 2012), когда постоянные и обильные дожди создают подходящую для развития головастиков среду, например пруды (Venâncio and Melo-Sampaio, 2010; Pinto et al., 2013). Длительный репродуктивный сезон вида обусловливает территориальность самцов, асинхронный приход самок к местам размножения, а также конкуренцию самцов за самок с физическим боем, включающий в себя физическое столкновение пар амфибий (рис. 1С; Souza 2009; Venâncio and Melo-Sampaio, 2010; Silva et al., 2020). Размножение сопровождает громкое кваканье и перемещению по растительности, что, возможно, повышает обнаруживаемость взрослых особей, в основном самцов, человеком.

Самцы подают голос поодиночке или небольшими скоплениями на растительности (ветвях кустарников и деревьев), вблизи ручьев, временных или постоянных водоемов на внутренней и опушечной частях леса, на высоте от 1 до 10 м над землей (Souza, 2009). Голос обычно начинается в сумерках и продолжается до рассвета, после чего самцы покидают места призыва и перемещаются в верхние дневные убежища (Souza, 2009). Ухаживания вида начинаются ночью в горных лесах с активного призыва самца с высокого дерева до прихода самки, после чего пара спускается в нижние дендрологические ярусы (Venâncio and Melo-Sampaio, 2010; Silva et al., 2020). Призыв самцов P. bicolor короткий, громкий, редкий и низкочастотный по сравнению с большинством древесных лягушек (Rodriguez and Duellman, 1994; Lima et al., 2012). При приближении самки самец забирается на её спину, образуя аксиллярный или цефалический амплексус (рис. 1А), в котором пара находится в течение нескольких минут, прежде чем самка переместится на подходящее место для яйцекладки (Venâncio and Melo-Sampaio, 2010). Место для яйцекладки выбирается на растительности над водной поверхностью (в среднем 70 см) или вблизи медленно текущего потока воды (рис. 1D), где спаривающаяся пара складывает листья (обычно листья деревьев), образуя камеру для защиты икры, что, вероятно, является стратегией против хищничества и обезвоживания (Venâncio and Melo-Sampaio, 2010; Lima et al., 2012; Pinto et al., 2013). Однако известно, что уровень хищничества на икре P. bicolor может достигать 61% (Neckel- Silveira and Washlewski, 2004).
Икра P. bicolor является одной из самых крупных среди древесных лягушек Амазонии и представляет собой студенистую массу с относительно крупными (в среднем 2,6 мм) икринками, количество которых варьирует от 241 до 1722 штук (Neckel-Oliveira and Wachlevski, 2004; Venâncio and Melo-Sampaio, 2010; Silva et al., 2020) (рис. 2В). После яйцекладки лецитотрофные яйца развиваются в головастиков в камерах листьев (Pinto et al, 2013) . Примерно через 2 недели из яиц выходят головастики с наружными жабрами (рис. 2А), которые опускаются на поверхность воды под местом яйцекладки и продолжают свое развитие с экзотрофным питанием до метаморфоза (рис. 2С) (Venâncio and Melo-Sampaio, 2010; Pinto et al., 2013).

1.3 Морфология и поведение
Phyllomedusa bicolor — относительно крупные древесные лягушки: взрослые самцы достигают 91-118 мм от рыла до брюшка, а самки — 106-119 мм (Souza, 2009; Lima et al., 2012; Duellman et al, 2016). Взрослые особи имеют темно-зеленую спину и белое до желто-белого или кремового брюхо (Lima et al., 2012). На нижних губах, груди и передних ногах имеются белые пятна с темными контурами, распределенные редко, но более густо с боку и сзади, темно-серая радужная оболочка, заметная железа, простирающаяся сзади глаз над барабанной перепонкой (Caramaschi and Cruz, 2002; Lima et al., 2012). Пальцы прозрачно-коричневые с крупными зелеными клейкими дисками, хотя на ногах и руках перепонок практически нет (Faivovich et al., 2010; Duellman et al., 2016). Обычно особи демонстрируют элегантную медленную походку по веткам и листьям, несмотря на способность к прыжкам (Caramaschi and Cruz, 2002). Виды Phyllomedusiae считаются фотогеничными и харизматичными животными, их часто называют «лягушками-плакатами» (Faivovich et al., 2010). Вероятно, из-за крупных размеров, широкого распространения и использования в шаманских ритуалах P. bicolor часто фигурирует в качестве символа амазонской фауны (Bernarde, 2012). Филломедузы активно выделяют кожные выделения и часто разносят их по телу задними и передними лапами (Blaylock et al., 1976).
По большинству характеристик головастики сходны с другими видами Phyllomedusa, т.е. являются экзотрофами, ленточниками и суспензорами (Altig and McDiarmid, 1999; Pinto et al., 2013). Головастик достигает общей длины до 50 мм (Souza, 2009). При жизни окраска спины и передней части тела оранжевая, брюшко серебристое, хвостовая мускулатура и хвост бледно-оранжевые, плавники полупрозрачные оранжевые, этого вида легко отличить от других (за исключением Phyllomedusa vaillantii) благодаря уникальному хвосту, присутствующему на всех стадиях развития, что подтверждает филогенетическую близость этих двух видов (Faivovich et al., 2010). Головастики Phyllomedusa bicolor имеют заметную окраску в дорсальной области, в то время как вентрально они считаются малозаметными. Вероятно, это позволяет им ориентироваться на воздушных или наземных хищников, оставаясь скрытными для подводных животных (Thibaudeau and Altig, 2012; Pinto and Amézquita, 2022). Они также предпочитают оставаться на поверхности воды, демонстрируя свою окраску: более крупные головастики демонстрируют такое поведение, в то время как мелкие особи обычно распределяются по толще воды, что усиливает антихищническую стратегию (Pinto and Amézquita, 2022). Имеются данные о токсичности головастиков P. bicolor для позвоночных: было показано, что после проглатывания они оказывают значительное воздействие на мышей, хотя наяды (личинки чернотелок) Odonata без труда их поедали. Поэтому вполне вероятно, что те же пептиды, которые секретируются взрослыми особями, присутствуют и во время роста личинок (Delfino et al., 1998; Pinto and Amézquita, 2022).
1.4 Географическое распространение и генетическая изменчивость
Этот вид имеет широкое распространение по сравнению с большинством видов амфибий, так как встречается в тропических дождевых лесах бассейна Амазонки в Боливии, Перу, Колумбии, Венесуэле, Гайане, Французская Гвиана, Суринам и, возможно, Эквадор (Ouboter and Jairam, 2012; Frost, 2021) (рис. 3). Однако большая часть его ареала обитания находится в Бразилии, где вид встречается в тропических лесах и даже в саванноподобных условиях всех северных штатов страны (Mota et al., 2020; Frost, 2021) (рис. 3).

Несмотря на крупные размеры тела, широкое распространение и медицинское значение, многие из немногих опубликованных последовательностей ДНК этого вида не относятся к какому-либо конкретному географическому региону, что объясняется тем, что они были получены из образцов, предназначенных для торговли домашними животными (Mota et al., 2020). Удивительно, но даже в филогенетических исследованиях, посвященных этому семейству, образцы P. bicolor отсутствуют (Wiens et al., 2005; Gomez-Mestre et al., 2008). Одно из исследований, посвященное пространственному распределению генетической изменчивости этого вида с использованием митохондриальных и ядерных маркеров, выявило две хорошо поддерживаемые монофилетические группы, которые разошлись примерно 5,16 млн. лет назад в двух географических мегарегионах — Восточной и Западной Амазонии; первая состоит из трех высокоструктурированных популяционных групп, распределенных в Гвианском и Бразильском щитах (Mota et al., 2020). Учитывая, что данное исследование было ограничено бразильскими местонахождениями и не содержало образцов из Суринама, типового местонахождения вида, вполне вероятно, что номинальный вид состоит как минимум из двух, а возможно, и более криптических видов (Mota et al., 2020; Frost, 2021).
1.5 Экологические взаимоотношения: Питание, хищничество и паразитизм
В природных условиях, по-видимому, нет данных о добыче P. bicolor. Другие филломедузы придерживаются генералистического рациона и считаются оппортунистическими хищниками, питающимися преимущественно арахнидами (пауками и клещами), колеоптилями и личинками лепидоптерообразных (Bertoluci, 2002; Lima et al., 2012).
Оказалось, что риск хищничества для ануранов меньше у видов с химической защитой или крупными размерами тела (Shine, 1979), что характерно для взрослых особей P. bicolor (Venâncio and Melo-Sampaio, 2010). Нет данных о том, что взрослые особи P. bicolor были добыты или найдены среди содержимого желудков таксонов, которые обычно охотятся на древесных лягушек, таких как змеи, птицы, летучие мыши или другие млекопитающие. Возможно, это связано с крупными размерами тела и загадочной окраской, которая помогает им маскироваться. Однако кожная секреция лягушки может играть важную роль в избегании хищников, поскольку она неприятна для них, возможно, вызывает регургитацию, изменяет сердечную деятельность или вызывает каталепсию (Sazima, 1974; Negri et al., 1992).
Некель-Оливейра и Вахлевский (2004) наблюдали, как икру P. bicolor хищнически поедали мухи, жуки и млекопитающие, а также сообщили, что хищничество икры — обычное явление для Phyllomedusa. Известно, что змеи хищничают на яйцах ануранов и могут быть причиной пропажи некоторых кладок (Warkentin, 1995; Martins and Oliveira, 1998; Neckel-Oliveira and Wachlevski, 2004). Некель-Оливейра и Вахлевский (2004) наблюдали следы от хищничества обезьян на кладки. Головастики Phyllomedusa bicolor были обнаружены среди содержимого желудка водной лягушки Pipa arrabali, и этот вид также был замечен активно охотящимся на головастиков, как только они падали из гнезда в водоем (Buchacher, 1993).
В качестве паразитов Phyllomedusidae отмечено всего девять видов нематод, а род Neocosmocercella был обнаружен только у филломедуз и, вероятно, является специфичным для этого семейства (Campião et al., 2014; Santos et al., 2017). Паразит Neocosmocercella fisherae был единственным видом, обнаруженным в толстом кишечнике особей P. bicolor (Santos et al., 2017). Также сообщалось о филяриевом паразите, обнаруженном вблизи легких P. bicolor, но без дальнейшей идентификации (Chai, 2015). Помимо хищничества мух на яйцах, для P. bicolor нет данных о миазе. Эндопаразиты, как кишечные, так и легочные, встречаются у P. bicolor чаще, чем эктопаразиты, что может отражать защиту от внешнего паразитизма, обеспечиваемую эпидермальными выделениями.
1.6 Сохранение и благополучие
Амазонские ануры с широким распространением, такие как P. bicolor, обычно отличаются криптическим таксономическим разнообразием, что означает, что виды, считающиеся не находящимися под угрозой, могут иметь неточный, а зачастую и недооцененный природоохранный статус (Gehara et al., 2014; Mota et al., 2020). Несмотря на широкое известное в настоящее время географическое распространение P. bicolor, этот вид не является многочисленным в фаунистических кадастрах и экологических исследованиях, а также рассматривается как менее многочисленный таксон в сообществах ануранов (Menin et al., 2008; Jairam, 2019). Несмотря на это, P. bicolor не отнесен к категории угрожаемых видов в приложении к Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (СИТЕС); это международное соглашение, цель которого — обеспечить, чтобы международная торговля дикими животными и растениями не угрожала выживанию видов (https://cites.org/eng). Согласно Красной книге угрожаемых видов Международного союза охраны природы (МСОП), которая отражает глобальный природоохранный статус вида (и срочно нуждается в обновлении), P. bicolor занесен в список видов, вызывающих наименьшую озабоченность, благодаря большому ареалу распространения и встречаемости на охраняемых территориях (МСОП, 2021).
Вышеупомянутый природоохранный статус P. bicolor может быть недооценен по следующим причинам: 1) Специфические требования к среде обитания, поскольку этот вид крайне нетерпим к антропогенным местообитаниям и обычно встречается в обширных и хорошо сохранившихся лесах (Tsuji-Nishikido and Menin, 2011; Lima et al., 2012). Их репродуктивное поведение отличается сложностью и требует подходящей наземной и водной (особенно ленточной) среды для завершения репродуктивного цикла; 2) Возможное криптическое разнообразие видов, встречающихся за пределами охраняемых территорий и/или с небольшим ареалом распространения (Mota et al., 2020); 3) Неизвестно популяционные эффекты, связанные с манипуляциями с особями для извлечения яда, которые иногда включают в себя использование электрического тока (Long et al., 2017; Liu et al., 2020; Chen et al., 2021) и сдерживающих устройств (Den Brave et al., 2014). Кроме того, неизвестно, как манипуляции и/или изъятие человеком взрослых особей из природы влияют на индивидуальное поведение, включая родительскую заботу и, как следствие, выживаемость потомства у P. bicolor; 4) Занижение количества жертв торговли животными. Как и в случае с любым другим запрещенным веществом, существует высокий риск ошибки идентификации, особенно из-за морфологического сходства видов Phyllomedusa или возможной фальсификации лиофилизированных экстрактов кожи (Lima and Labate, 2007; Silva et al., 2019). Например, известно, что в столичном регионе Манаус обитает не менее четырех видов Phyllomedusa, встречающихся в симпатрии (в одном и том же месте) и синтопии (в сходных микроместообитаниях) (Lima et al., 2012), что может привести к неправильной идентификации видов и извлечению кожных выделений нецелевых видов.
Phyllomedusa bicolor распространена в основном в Бразилии, стране с ограничительным законодательством, запрещающим использование и рекламу камбо в медицинских целях без должного разъяснения рисков и отсутствия доказательной базы для лечения (ANVISA, 2004). Кроме того, существуют жесткие ограничения в отношении доступа к диким животным и их продуктам даже для научных исследователей (IBAMA/ICMBIO). Несмотря на это, основные центры, где применяется секреция, находятся за пределами ареала распространения вида (Iakp, 2022; Thompson and Williams, 2022), а информация о том, как особи и секреция попадают в эти места, отсутствует, поскольку в центрах применения редко указывается точное происхождение и таксономическая точность отобранных особей. В 2013 году Региональное управление федеральной полиции бразильского штата Акре разработало методику химической идентификации «вакцины против жаб» (vacina do sapo) и пресечения незаконного оборота и контрабанды, однако информация о текущем применении такой методики отсутствует.
2 Ядовитая секреция Phyllomedusa bicolor
2.1 Строение кожи Phyllomedusa bicolor
Кожа амфибий — тонкий, проницаемый и гибкий покров, обладающий высокой сосудистой активностью и отвечающий за поглощение воды, дыхание, терморегуляцию, осморегуляцию, физическую и химическую защиту от хищников и высыхания (Wager, 1986; Clarke, 1997; Larsen and Ramløv, 2013). Она состоит из слоев эпидермиса и дермы, и на всей эпидермальной поверхности лягушек имеется бесчисленное количество желез (рис. 4), которые постоянно вырабатывают и выделяют небольшие количества яда (Jared et al., 2009; Govender et al., 2012). Среди амфибий дендробиальные лягушки, такие как древесные лягушки, проводят большую часть жизни во влажной среде, где в результате чего повышается подверженность особей инфекционным заболеваниям (Liu et al., 2021). Кожа ануранов приспосабливается к различным воздействиям окружающей среды в местах обитания вида, что обусловливает большое морфофункциональное разнообразие (Toledo and Jared, 1993; Barra and Simmaco, 1995). Их выделения играют важную роль в обеспечении кожного дыхания, размножения, защиты от хищников, а также защиты от высыхания и размножения микроорганизмов (Toledo and Jared, 1995).
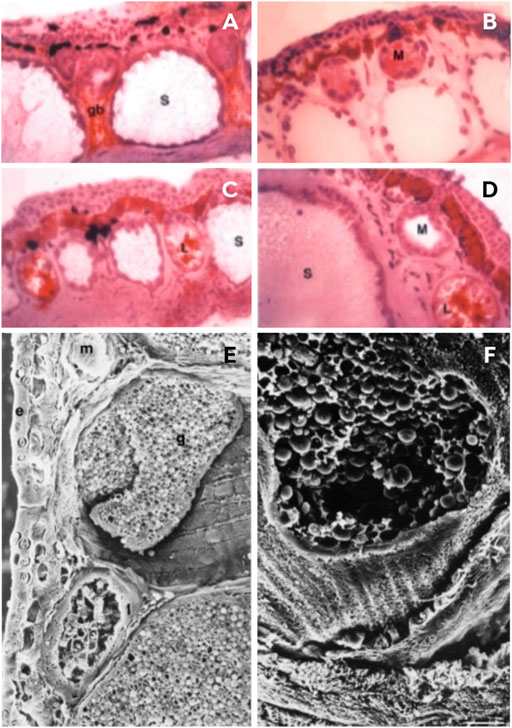
Lacombe et al. (2000) провели ультраструктурное исследование, в котором охарактеризовали кожу видов Phyllomedusa по размеру и активности, и представили профиль кожных желез, состоящий из трех типов кожных желез: липидных, слизистых и серозных (рис. 4). Эти железы расположены глубоко в коже и подкожной соединительной ткани, хотя серозные железы крупнее и лежат глубже остальных (Lacombe et al., 2000). Липидные железы расположены в основном в дорсальной и дорсолатеральной областях и отвечают за герметичность кожи, а также предотвращают потерю воды при высыхании (Lacombe et al., 2000; Castanho and De Luca, 2001). Слизистые железы расположены в основном в вентральной области и практически отсутствуют на спине. Они вырабатывают слизь для обеспечения таких физиологических функций кожного покрова, как дыхание, размножение, защита и терморегуляция (Toledo and Jared, 1995; Lacombe et al., 2000). Серозные железы являются самыми крупными и распределены по всему телу, однако они выделяются в дорсолатеральной области и за глазами, образуя околоушные железы (Lacombe et al., 2000). Эти железы отвечают в основном за пассивную защиту и развиваются первыми, хотя проток железы удлиняется в зависимости от созревания эпидермиса и открывается на поверхность кожи по достижении метаморфоза (Toledo and Jared, 1995; Lacombe et al., 2000). По данным Lacombe et al. (2000), существует два класса серозных желез — I и II типа. Первый тип имеет слабо развитый гладкий эндоплазматический ретикулум, который подразделяется на два подтипа — Ia и Ib. Тип Ia характеризуется наличием плотных гранул, в которых осуществляется биосинтез белковых продуктов, предназначенных для экзоцитоза (рис. 4H), и задействован как шероховатый эндоплазматический ретикулум, так и аппарат Гольджи (Delfino, 1991). В серозном секрете типа Ib видны везикулы, несущие люцентный материал, который подвергается созреванию без конденсации (Toledo and Jared, 1995). Тип II характерен для P. bicolor, поскольку они имеют хорошо развитый, гладкий эндоплазматический ретикулум, который, вероятно, играет роль в биосинтезе пептидов (Blaylock et al., 1976; Lacombe et al., 2000). Сначала эти пептиды синтезируются как препропептиды, затем, для их созревания, происходит удаление пептидного сигнала и кислого пропика, после чего зрелые пептиды накапливаются в круглых гранулах (Lacombe et al., 2000; Nicolas and El Amri, 2009; Calderon et al., 2011).
2.2 Состав кожного секрета Phyllomedusa bicolor
Амфибии обитают в среде, богатой микроорганизмами, особенно древесные лягушки, которые основную часть жизни живут в очень влажной среде. В результате они становятся более восприимчивыми к патогенам и, чтобы защититься, вырабатывают мощные антимикробные пептиды (Daly, 1995; Govender et al., 2012; Liu et al., 2021). Система врожденного иммунитета несет основную ответственность за выживание особи, при этом секреция, вырабатываемая железами ануранов, подавляет развитие заболеваний и является токсичной для других животных, что в некоторых случаях может предотвратить хищничество (Nicolas and Mor, 1995; Kimbrell and Beutler, 2001; Toledo et al., 2011). Выделение кожного секрета считается очень эффективным защитным механизмом, поскольку он обеспечивает невкусные характеристики и/или токсичность в отношении потенциальных хищников (Bevins and Zasloff, 1990; Daly, 1995; Williams et al., 2000; Rodriguez et al., 2011; Saporito et al., 2012; Hantak et al., 2013). Среди химических веществ, ответственных за защиту ануранов, основную роль играют пептиды, в то время как алкалоиды встречаются реже и выделяются некоторыми видами в тело (Saporito et al., 2009; Jeckel et al., 2015). Известно, что пептиды в основном действуют против патогенных микроорганизмов, однако исследования показали, что они также могут выполнять функции, связанные с взаимодействием между видами, например брадикининоподобные пептиды, выделяемые Bombina orientalis (Bombinatoridae), которые, как известно, способствуют процессу интоксикации хищников (Rödel et al., 2013; Raaymakers et al., 2018). Несмотря на то что кожные выделения Phyllomedusa bicolor широко известны как богатый источник биологически активных пептидов, до сих пор не проводились исследования, направленные на выяснение того, обладают ли вещества, выделяемые в кожных выделениях, токсичностью в отношении хищников (Bevins and Zasloff, 1990; Daly, 1995).
Все виды Phyllomedusa являются активными продуцентами пептидов, среди которых наиболее представительным семейством являются дермасептины (Erspamer et al., 1985). С момента первой записи о выделении пептида из этого рода (датируемой 1966 годом) было описано более 277 пептидов с уникальными последовательностями (Calderon et al., 2011). Большинство охарактеризованных к настоящему времени биологически активных пептидов имеют потенциальное применение в медицине: филлокаэрулеины обладают гипотензивными свойствами, тахикинины и филлокинины — вазодилататорами, дерморфины и дельторфины — опиатоподобными свойствами, аденорегулин — антибиотическими свойствами (Daly et al., 1992; Mor et al., 1994a; Lacombe et al., 2000; Hesselink and Sacerdote, 2019). В соответствии с основной активностью эти пептиды делятся на группы: антимикробные пептиды, пептиды, действующие на гладкую мускулатуру, и пептиды, действующие на нервную систему (Erspamer et al., 1981). Благодаря многочисленным биологическим свойствам этих веществ и сходству с аминокислотными последовательностями нейропептидов и гормонов млекопитающих, многие из них вызывают интерес с медицинской и фармакологической точек зрения, например, для получения новых лекарственных препаратов (Lazarus et al., 1999; Basir et al., 2000; Chen et al., 2002; Doyle et al., 2002; Severini et al., 2002; Conceição et al., 2006).
Яд P. bicolor используется туземцами в традиционных шаманских ритуалах на юго-западе Амазонки и, по некоторым данным, применяется как стимулирующее средство, а также в лечебных целях, хотя убедительных научных доказательств его эффективности нет (Erspamer-Falconieri et al., 1970; Majic et al., 2021). К настоящему времени из кожных выделений P. bicolor охарактеризованы только пептиды, из которых определено 28 различных последовательностей, причем большинство из них относится к семействам дермасептина, дельторфина и деморфина (Thompson and Williams, 2022).
2.2.1 Дермасептины
Дермасептины представляют собой суперсемейство антимикробных катионных пептидов, вырабатываемых в коже Phyllomedusidae. Известно, что они генетически родственны, сходны по сигналам и последовательностям предшественников, но впоследствии диверсифицировались и дали начало структурно различным пептидным подсемействам (Nicolas and El Amri, 2009). Дермасептины образуются из предшественников, имеющих высококонсервативный N-концевой препроцессор (Nicolas and Amiche, 2006). Дермасептины имеют родственные структуры, отличающиеся длиной последовательности, а также различиями в аминокислотных последовательностях. Первым охарактеризованным дермасептином был S1 (DRS1), присутствующий в коже P. sauvagii (Mor et al., 1991), а для P. bicolor выделенные/охарактеризованные в его коже дермасептины пока относятся к серии B (B и B1-B6), G (G3) и другим классам дермасептиноподобных пептидов (табл. 1). За исключением DRS-S4, все дермасептины обладают низкой или нулевой токсичностью для клеток млекопитающих (Ladram and Nicolas, 2016). Избирательность и способность дестабилизировать плазматическую мембрану — наиболее значимая характеристика антимикробного действия дермасептинов. Это семейство антимикробных пептидов в основном отвечает за противоинфекционный барьер пассивной защиты, в котором механизм киллинга действует быстро, разрушая плазматическую мембрану патогена за несколько минут, и вряд ли может вызвать антибиотикорезистентность (Shai, 1995; Feder et al., 2000).
2.2.1.1 Дермасептины stricto sensu
По химическим характеристикам большинство дермасептинов stricto sensu имеют амидированный С-концевой остаток и аминокислоту лейцин в положении 2 первичной структуры, за исключением дермасептинов В и В1, у которых в этом же положении находятся валин и метионин соответственно. Кроме того, они имеют остаток триптофана в положении 3 первичной структуры (Daly et al., 1992; Mor et al., 1994a; Charpentier et al., 1998; Fleury et al., 1998; König et al., 2012) (табл. 1). Большинство пептидов из P. bicolor содержат посттрансляционно модифицированные аминокислотные остатки, которые играют важную роль в активности и рецепторной специфичности (Conibear, 2020). Кроме того, некоторые модификации могут обеспечивать кросс-сопряжение специфических аминокислот, что позволяет получать производные с повышенной биостабильностью. Кроме того, включение таких модификаций может обеспечить устойчивость к протеолитической деградации, особенно для антимикробных пептидов (Tiwari, 2019). Поскольку они являются α-спиральными, катионными и амфипатическими, то действуют непосредственно на клеточную мембрану микроорганизмов, дестабилизируя фосфолипидные основы и препятствуя входу и выходу веществ, что приводит к лизису клетки (Nicolas and Amiche, 2006). Для P. bicolor известно семь пептидных последовательностей дермасептина stricto sensu (все из серии В), а именно: дермасептин В (DRS-B), дермасептин В1 (DRS-B1), дермасептин В2 (DRS-B2), дермасептин В3 (DRS-B3), дермасептин В4 (DRS-B4), дермасептин В5 (DRS-B5) и дермасептин В6 (DRS-B6). Количество остатков дермасептина серии В, строго говоря, обнаруженных в P. bicolor, варьирует от 24 до 33 аминокислот (Charpentier et al., 1998; Fleury, 1998; König et al., 2012) (табл. 1).
Пептид DRS-B был впервые выделен Mor et al. (1994a) и представлял собой амид пептида с 27 остатками в последовательности, в которой остатки лизина чередуются гидрофобными и гидрофильными участками. DRS-B демонстрирует 78% сходство с дермасептином S, отличаясь от него лишь рядом делеций и замен (табл. 1). Mor et al. (1994a) первыми испытали ДРС-В против патогенных грибов и бактерий. Были определены минимальные ингибирующие концентрации (МИК) против восьми видов грибов и пяти видов бактерий. Антимикробный потенциал ДРС-Б проявился в отношении грибов Cryptococcus neoformans, Candida albicans, Microsporum canis, Tricophyton rubrum, T. mentagrophytes, Arthroderma simii, Aspergillus fumigatus и A. niger и бактерий Aeromonas caviae, Escherichia coli, Enterococcus faecalis и Nocardia brasiliensis, поскольку значения МИК составляли от 40 до 60 мкМ (Mor et al., 1994a).
Аналогичным образом все остальные дермасептины stricto sensu (DRS-B1 — DRS-B6) проявили активность в отношении микроорганизмов. DRSB1 имеет в своей первичной структуре 31 аминокислотный остаток и, в отличие от других дермасептинов stricto sensu, имеет остаток метионина в положении 2 первичной последовательности. С другой стороны, DRS-B2 имеет в своей структуре 33 аминокислотных остатка. В отличие от других дермасептинов, Van Zoggel et al. (2012) показали, что DRS-B2 подавляет рост аденокарциномы предстательной железы и некоторых других клеточных линий. DRS-B3, напротив, имеет в своей первичной структуре 28 аминокислотных остатков, но отличается от остальных тем, что не имеет модификаций на С-концевом остатке. Пептид DRS-B4, как и DRS-B3, имеет такое же количество аминокислотных остатков, однако в нем присутствует модифицированная С-концевая группа (Van Zoggel et al., 2012) (табл. 1). И DRS-B3, и DRS-B4 проявили антибактериальную активность с МИК в диапазоне от 1,3 до 11,6 мкМ (Van Zoggel et al., 2012) (табл. 1). Кроме того, DRS-B4 проявил антидиабетическую активность: обработка этим пептидом глюкозореактивных клеток BRIN-BD11 приводила к стимуляции высвобождения инсулина (Marenah et al., 2004).
Пептид DRS-B5, состоящий из 25 аминокислотных остатков, пока не оценивался с точки зрения биологического потенциала, но предполагается его антимикробный потенциал, как и у других дермасептинов (Charpentier et al., 1998). Аналогичным образом DRS-B6, первичная последовательность которого состоит из 24 аминокислотных остатков, также не изучался на предмет фармакологического потенциала. DRS-B6, как и DRS-B1, были связаны с механизмами химической защиты P. bicolor, поскольку они секретируются серозными железами, расположенными в голенях (König et al., 2012). Наконец, синтетический дермасептин, названный DRS-G3, был идентифицирован с помощью метода кДНК в P. bicolor и синтезирован с использованием 9-фторенилметоксикарбонил-защищенных (FMOC) аминокислот (Fleury et al., 1998). В первичной структуре этого пептида 30 остатков с консервативным С-концом. Можно отметить, что он имеет три аминокислотных остатка (Ala, Leu и Trp), которые характерны для большинства других дермасептинов, присутствующих в P. bicolor. Fleury et al. (1998) проанализировали биологическую активность DRS-G3 и отметили, что, как и известные дермасептины, он также обладает антимикробной активностью в отношении бактерий. Авторы, помимо МИК, проверили минимальную летальную концентрацию (МЛК), необходимую для уничтожения бактерий, таких как Mollicutes, Firmicutes и Gracilicutes, и обнаружили, что МИК варьирует от 3,0 до 6,25 мкМ, а МЛК — от 6,25 до 100 мкМ (табл. 1).
2.2.1.2 Другие дермасептины
Аденорегулин представляет собой поликатионный пептид, состоящий из 33 аминокислотных остатков в первичной структуре (табл. 1). Это вещество имеет α-спиральную структуру с лизинами, расположенными на внешней стороне (Daly et al., 1992). По сравнению с дермасептинами аденорегулин имеет ряд замен в аминокислотных остатках и не имеет модификаций на С-конце. Cao et al. (2005) получили рекомбинантное производное аденорегулина путем гетерологичной экспрессии в E. coli. Это производное отличается от исходной молекулы наличием дополнительного амидированного остатка глутамина в С-концевой части, что увеличило силу его антимикробной активности в отношении Tritirachium album и S. cerevisiae.
Помимо антимикробных свойств, аденорегулин в низких концентрациях обладает противоопухолевыми и ангиостатическими свойствами. Интересно, что противоопухолевая активность этой молекулы связана с некрозом клеток (Santos et al., 2017). Кроме того, упоминается потенциал в регуляции клеточного метаболизма, поскольку этот пептид может связываться с рецепторами аденозина А1, что делает его потенциальной молекулой для лечения депрессии, болезни Альцгеймера и болезни Паркинсона. (Cao et al., 2005).
Дермотоксин — пептид, полученный из тех же предшественников семейства дермасептина, представляет собой катионный и амфипатический пептид с 32 аминокислотными остатками в первичной структуре и не имеет модификаций на С-конце (Amiche et al., 2000) (табл. 1). У P. bicolor отмечено, что дермотоксин экспрессируется как в коже, кишечнике, так и в головном мозге. Дерматоксин, как и другие представители семейства дермасептинов, обладает эффективной антимикробной активностью широкого спектра действия. Amiche et al. (2000), испытав дерматоксин, обнаружили антибактериальную активность с МИК от 6,25 до 100 мкМ в отношении грамположительных бактерий Bacillus megaterium и Corynebacterium glutamicum, бактерий без клеточных стенок A. laidlawii и Spiroplasma melliferum, а также грамотрицательной бактерии Sinorhizobium meliloti.
Филлоксин также происходит из группы предшественников дермасептина и имеет в своей первичной структуре 19 аминокислотных остатков без посттрансляционных модификаций (Pierre et al., 2000) (табл. 1). Pierre et al. (2000) обнаружили антимикробную активность в отношении некоторых штаммов бактерий, однако с меньшей способностью к ингибированию, чем другие дермасептины. Филлоксин оказался перспективным в отношении A. laidlawii, Bacillus megaterium, Spiroplasma melliferum, Escherichia coli и Micrococcus luteus. Что касается других фармакологических свойств, то информация об их антимикробной активности отсутствует.
Пептид филлосептин-B2 был впервые выделен у Phyllomedusidae из кожного секрета P. sauvagii (Zhang et al., 2010), а позднее его аналог был охарактеризован у P. bicolor путем клонирования кДНК (König et al., 2012). Поскольку предшественники пептида в P. bicolor ранее не были идентифицированы, а ортология с родственными последовательностями P. sauvagii и P. hypochondrialis не гарантирована, пептид был обозначен как филлосептин-B2, хотя дальнейшие исследования по этому вопросу должны быть проведены. Антибиотические свойства лизиса плазматической мембраны пептиды проявляют в виде нескольких пузырьковых образований, разрушающих мембрану (Leite et al., 2005). Спектр антимикробной активности включает грамотрицательные бактерии, например Acinetobacter calcoaceticus, E. coli и Pseudomonas aeruginosa (Leite et al., 2005; Resende et al., 2008); грамположительные бактерии, например Enterococcus faecalis, Klebsiella peneumoniae, Staphylococcus aureus и Streptococcus agalactiae; грибы: C. albicans (Resende et al., 2008); и простейшие, например, Leishmania amazonensis (промастиготы) (Kückelhaus et al., 2009), Plasmodium falciparum (кольца, трофозоиты и шизонты) (Kückelhaus et al., 2009) и Trypanosoma cruzi (трипомастиготы) (Leite et al., 2005). Однако токсическое действие на клетки маммалл наблюдалось, но только в экстремально высоких концентрациях (Kückelhaus et al., 2007; Kückelhaus et al., 2009), а влияние на клетки крови было незначительным (Leite et al., 2005).
2.2.2 Панкреатические полипептиды (ПП)
Панкреатические полипептиды (ПП) были выделены при получении инсулина и являются первыми из семейства ПП. Другим пептидом этого семейства является тирозиновый пептид (PYY), характерной особенностью которого является С-концевой амидированный остаток тирозина (Grandt et al., 1994). У ануранов известны производные PP, особенно у вида Xenopus laevis, функция которых в организме еще недостаточно изучена (Kim et al., 2001).
У вида P. bicolor единственным охарактеризованным на сегодняшний день ПП является кожный пептид тирозина (SPYY), выделенный Mor et al. (1994a). Это вещество имеет первичную структуру, состоящую из 36 аминокислотных остатков, в которой С-концевой остаток амидирован (табл. 1). Функция этого пептида у P. bicolor пока неясна, однако была проверена его ингибирующая активность в отношении альфа-меланоцитстимулирующего гормона (α-MSH), и удалось продемонстрировать, что in vitro SPYY подавляет секрецию α-MSH (Mor et al., 1994b). Антимикробная активность этих пептидов распространяется на штаммы грамотрицательных бактерий (например, Aerornonas caviae и E. coli), грамположительных бактерий (например, Enterococcus faecalis и Nocardia brasiliensis), грибов (например, Arthroderma simii, Aspergillus fumigatus, A. niger, Microsporum canis) и простейших (например, промастиготы Leishmania major) (Calderon et al., 2011).
2.2.3 Кальцитонины
Это семейство включает пептиды с 32-53 аминокислотными остатками (Hay et al., 2017), которые признаны гормонами, способными быстро снижать уровень циркулирующего кальция в организме (Naot et al., 2018). Снижение происходит за счет ингибирования эффлюкса кальция из костей, поскольку пептиды этого класса выступают в качестве мощных ингибиторов костной резорбции (Naot et al., 2018). Известны и другие вещества этого семейства, такие как амилин, адренорегулин (Hay et al., 2017).
Кожный кальцитонин-ген-связанный пептид (SCGRP) был выделен из кожи особей P. bicolor Сеоном и др. (2000). Первичная структура этого пептида состоит из 37 аминокислотных остатков, причем С-концевой остаток амидирован, а между цистеинами в положениях два и семь имеется дисульфидный мостик (табл. 1). Наличие дисульфидной связи может быть связано со стабилизацией всей трехмерной структуры SCGRP, что может иметь решающее значение для гормональной функции (Góngora-Benítez et al., 2014). Исследователи, открывшие этот пептид, провели анализ связывания с рецептором кальцитонина и заметили, что SCGRP отличается от всех других представителей кальцитонинов как минимум в девяти позициях. Как и другие пептиды этого семейства, он показал конкурентное ингибирование и высокую специфическую аффинность связывания с кальцитониновыми рецепторами в мозге крыс (Seon et al., 2000).
2.2.4 Кортикотропин-рилизинг гормоноподобный пептид
Совагин — единственный пептид из семейства кортикотропин-рилизинг-гормоноподобных пептидов, идентифицированный у P. bicolor (Erspamer et al., 1993). Он имеет 40 аминокислотных остатков, в которых С-концевой остаток амидирован, а также наличие пироглутаминовой кислоты в качестве N-концевого остатка (табл. 1). Это вещество также связано с некоторыми эффектами камбо, а также обладает гипотензивной и антидиуретической активностью у крыс (Erspamer et al., 1981). Многообещающие результаты были получены при оценке действия совагина на состояние гиперпролактинемии, когда 500 мкг совагина, введенные путем подкожной инъекции, вызывали немедленное снижение повышенного уровня пролактина в сыворотке крови до нормальных значений в течение 5 ч (Erspamer et al., 1981).
Этот пептид имитирует функциональные последствия стрессового воздействия и действует как кортикотропин-рилизинг фактор (КРФ) (Lovejoy and Balment, 1999; Hesselink and Sacerdote, 2019). Он имеет два подтипа CRF-рецепторов (CRFR), которые связываются с CRFR1 и CRFR2, и опосредует тревогу благодаря агонизму CRFR2 (Eckart et al., 1999; Hesselink and Sacerdote, 2019). Эффекты в исследованиях на животных моделях связаны с последовательностью интенсивного, длительного гипотензивного действия, за которым следуют тахикардия, антидиурез, снижение скорости гломерулярной фильтрации (СКФ) и увеличение канальцевой реабсорбции Na+ (Montecucchi et al., 1979; Erspamer et al., 1981; Heinrichs and Taché, 2001). Кроме того, происходит снижение температуры тела, обусловленное механизмом, опосредованным дофаминовыми рецепторами D2 (Hesselink and Sacerdote, 2019). В моделях на собаках он проявлял различные эффекты, такие как длительное увеличение кровотока (Lovejoy and Balment, 1999).
2.2.5 Тахикинины
Тахикинины — это семейство пептидов, присутствующих у амфибий и млекопитающих и проявляющих нейротрансмиттерную активность (Steinhoff et al., 2014). Они экспрессируются во всей нервной и иммунной системе, регулируют чрезвычайно разнообразные физиологические процессы, а также участвуют в важных патологических состояниях (Steinhoff et al., 2014). У ануранов наличие тахикининов впервые было описано Шеффером и Габриэлем (2005), которые обнаружили присутствие этого пептида в сетчатке глаза особи рода Pelobates. У P. bicolor единственным известным на сегодняшний день тахикинином является филломедузин.
Филломедузин представляет собой декапептид, который впервые был охарактеризован Анастази и Эспармером (1970). Интересно, что он имеет пироглутаминовую аминокислоту в качестве N-концевого остатка, а также амидирован в С-конце (табл. 1). Что касается его биологической активности, то известно, что этот пептид способствует сокращению гладких мышц, снижает артериальное давление при интенсивной компенсаторной тахикардии, а также способствует высвобождению антидиуретических гормонов с последующим антидиурезом (Anastasi and Erspamer-Falconieri, 1970; Erspamer-Falconieri et al., 1970). Кроме того, было показано, что у крыс этот пептид увеличивает выброс в плазму кортикостерона, катехоламинов и глюкозы, а также высвобождение β-эндорфинов (Mathison and Solomos, 1985).
2.2.6 Брадикинин
Пептиды, связанные с брадикинином (BRPs), представляют собой широко распространенный класс кожных пептидов амфибий, которые имитируют действие гормонов брадикинина позвоночных и секретируются в так называемых гранулярных или серозных железах (Raaymakers et al., 2018). Поскольку виды Phyllomedusa отличаются большим разнообразием пептидов, содержащихся в их теле, можно предположить, что они могут усиливать поглощение брадикинина эпителием, ускоряя тем самым процесс интоксикации хищника, как это было показано Raaymakers et al. (2018) для Bombina orientalis. Что касается филлокинина P. bicolor, то это единственное вещество данного семейства, идентифицированное из его надкопытника, и использование БРП в защитных мероприятиях против хищников неизвестно (König et al., 2012).
Филлокинин имеет 11 аминокислотных остатков в своей первичной структуре, а в качестве С-концевого остатка имеет сульфатированную аминокислоту тирозин (Anastasi et al., 1966) (табл. 1). В фармакологических исследованиях на собаках было показано, что этот пептид сильнее, чем брадикинин, снижает артериальное давление; при отсутствии посттрансляционной модификации эта активность значительно снижается (Anastasi et al., 1966). Что касается ритуала камбо, то известно, что этот пептид вызывает некоторые симптомы, такие как учащение сердцебиения, приливы жара и покраснение кожи (Thompson and Williams, 2022).
2.2.7 Дельторфины
Дельторфины — это небольшое семейство короткоцепочечных пептидов, в первичной структуре которых насчитывается от шести до семи аминокислотных остатков (Erspamer et al., 1989). У ануранов это семейство пептидов ограничено родом Phyllomedusa (в основном P. bicolor и P. sauvageii) (Erspamer et al., 1989). Пептиды этой серии отличаются высоким сродством и селективностью к δ-опиоидным рецепторам, что делает их перспективными анальгетиками, поскольку по силе действия они в 4000 раз превосходят морфин и в 40 раз — эндогенные β-эндорфиновые рецепторы (Esparmer et al., 1989). Эти результаты были подтверждены исследованиями in vivo, показавшими, что дельторфины обладают высокой скоростью проникновения через гематоэнцефалический барьер (Fiori et al., 1997). Небольшой размер дельторфинов (также дермофинов) может быть связан с их функциями и/или биологической пользой для человека. В частности, небольшие пептиды, содержащиеся в P. bicolor, являются наиболее сильными анальгетическими соединениями, однако связь между их размером и биологической пользой остается неясной.
Первый дельторфин был обнаружен в P. sauvageii (Montecucchi et al., 1981a) и послужил основой для обозначения аналогичных пептидных последовательностей, описанных позднее в P. bicolor. Интересной особенностью этой первой находки было то, что в этом пептиде второй аминокислотный остаток (метионин) имеет инвертированную стереохимию, представляя собой D-конфигурацию (табл. 1).
В 1989 г. Эрспамер и др. (1989) выделили из кожного секрета P. bicolor два гептапептидных аналога, которые были названы [D-Ala2]-дельторфин I и [D-Ala2]-дельторфин II. В отличие от дельторфина, они имеют замену метионина на аланин в положении 2 первичной последовательности и два остатка валина подряд на С-конце. Еще одним отличием этих пептидных аналогов является замена аминокислотного остатка в положении 4 первичной структуры, причем у [D-Ala2]-дельторфина I это глутаминовая кислота, а у [D-Ala2]-дельторфина II — аспаргиновая кислота (Erspamer et al., 1989). В том же исследовании они продемонстрировали сродство этих пептидов к опиоидным рецепторам, которое в 10-200 раз выше, чем у синтетического энкефалина.
2.2.8 Дермофины
Дерморфины являются опиоидными пептидами и впервые были выделены из P. sauvagii (Montecucchi et al., 1981a), P. burneisteri и P. rohdei (Montecucchi et al., 1981b; Broccardo et al., 1981). Как и дельторфины, они обладают высоким сродством и селективностью к опиоидным рецепторам и, как было показано, вызывают анальгезию как у человека, так и у животных (Negri, 1992). Однако сродство и селективность дерморфинов относятся к µ-рецепторам, в то время как дельторфины являются специфическими агонистами δ-рецепторов (Richter et al., 1990). По сравнению с морфином дерморфин имеет потенциально более благоприятный профиль побочных эффектов, так как является селективным агонистом на корректоре Нидса к ϻ-опиоидным рецепторам (MOR), но не обладает соответствующим сродством к ƙ-опиоидным рецепторам (KOR) (Pescatore et al., 2015; Hesselink et al., 2018b). В доклинических исследованиях анальгетической активности при интрацеребровентрикулярном применении пептид дерморфин показал лучшую эффективность, чем морфин, и продемонстрировал более продолжительный и мощный эффект (Broccardo et al., 1981). Пептиды этого класса, как правило, воздействуют на центральную нервную систему, вызывая угнетение дыхания, а также высокую зависимость от постоянного потребления в ритуале камбо (Nakata et al., 1986).
У P. bicolor описаны три новых вещества, аналогичных дерморфину, кДНК которых были клонированы непосредственно из кожи. Они были названы Рихтером и др. (1990) новым дерморфином I, новым дерморфином II и новым дерморфином III. Новые аналоги I и III представлены в виде гептапептидов, а новый аналог II — пятью аминокислотными остатками в своей последовательности (табл. 1). В отличие от дермофина, они имеют консервативный С-концевой участок и, хотя по всей цепи наблюдаются делеции и замены, можно отметить, что они имеют сходную последовательность в трех аминокислотах N-концевой части (Tyr-D-Ala-Phe).
После химического синтеза этих предсказанных пептидов была проведена их оценка в отношении опиоидных рецепторов, в которой активность проявил только пентапептид, но с IC50 (половиной максимальной ингибирующей концентрации), которая была в 10 раз ниже, чем у дерморфина (Richter et al., 1990). Впоследствии из кожи P. bicolor были выделены два новых гептапептида класса дерморфинов (Mignogna et al., 1992). Первый из них называется [Lys7]-дерморфин, который отличается от модельного пептида класса наличием лизина в седьмом положении (Mignogna et al., 1992). С другой стороны, пептид [Trp4,Asn7]-дерморфин имеет две модификации — аминокислотный остаток триптофана в положении 4 и аспарагина в положении 7 (табл. 1). В положении 2 эти два аналога имеют аминокислоту аланин в L-конфигурации, а также консервативный С-концевой участок (Mignogna et al., 1992). Как и другие пептиды подобного типа, эти пептиды также проявили сродство к опиоидным рецепторам, однако оценка показала, что амидированные производные в 2-4 раза сильнее, чем варианты без посттрансляционной модификации (Mignogna et al., 1992).
2.2.9 Каэрулеины
Это семейство ограничено двумя пептидами, причем у вида P. bicolor охарактеризован только неапептид филлокаэрулеин (Anastasi et al., 1969). Он является аналогом пептида caerulein и отличается от него тем, что имеет на одну аминокислоту меньше, но сохраняет все характеристики своего сородича, такие как состав и две посттрансляционные модификации: тирозин с сульфатом и С-концевая амидирована (Anastasi et al., 1969). Еще одним необычным аспектом этого пептида является наличие в качестве N-концевого остатка аминокислоты пироглутаминовой кислоты. Филлокаэрулеин считается наиболее многочисленным пептидом, который вместе с совагином отвечает за желудочно-кишечные эффекты, наблюдаемые в ритуале камбо, в частности, тошноту, рвоту и снижение артериального давления (Thompson and Williams, 2022).
2.2.10 Бомбезины
Пептиды этого класса состоят из девяти аминокислот и имеют пироглутаминовую кислоту в качестве N-концевого остатка, а также амидированный С-концевой остаток (табл. 1). Их количество в P. bicolor ограничено следовыми количествами, среди которых выделяется пептид филлолиторин (Erspamer et al., 1985). Что касается их фармакологии, то они, как правило, стимулируют секрецию желудочных кислот и увеличивают секрецию поджелудочной железы, поскольку связаны с гастрин-рилизинг пептидом. Кроме того, они были описаны как пептиды, способные подавлять потребление алкоголя и пищи после оценки in vivo у цыплят (Cline et al., 2010). Следует отметить, что существуют противоречивые данные о наличии бомбезинов в секретах P. bicolor (Thompson and Williams, 2022). Однако исследования также связывают присутствие филлолиторина с ознобом, наблюдаемым у людей во время ритуала камбо (Esakov et al., 1990).
3 Ритуал Камбо: от традиционного использования к применению в крупных городских центрах
Жабы, лягушки и лягушки-древолазы — животные, которые часто встречаются в фольклоре и культуре практически повсеместно и ассоциируются с дождями и плодородием (Lima and Labate, 2007). Вакцина камбо, капум или жабья вакцина («vacina do sapo», на португальском языке) является ритуалом очищения и связана с ритуалами исцеления в тропических лесах Амазонки и городских центрах по всему миру (Rudgley, 1993; Lima and Labate, 2007; Haddad and Martins, 2020). Ритуал камбо традиционно проводится шаманами для очищения организма, увеличения физической силы и сексуальной выносливости, а также для защиты от «панемы» (невезения и слабости) (Balée, 2004; Lima, 2005; Bernarde and Santos, 2009; Labate and Lima, 2014). Он традиционно используется группами местных охотников на юго-западе Амазонии, среди которых яванауа, каксинава, матсес, майоруна, яванава и особенно катукина (Daly et al., 1992; Hesselink, 2018a). Носители языка пано также применяют kambô к собакам, которых они используют во время охоты (Souza et al., 2002). В этнологической литературе имеются сведения об использовании камбо более чем у 15 туземных групп, принадлежащих к пяти языковым семьям (пано, аруак, арава, тикуна и тупи-гуарани), проживающих в Боливии, Бразилии, Колумбии, Перу, Французской Гвиане, Суринаме и Венесуэле (Lima, 2000; Lima, 2005; Lima, 2008; Lima, 2009; Lima, 2012; Lima, 2014a; b; Ribeiro, 2017).
В этом ритуале шаман-целитель прижигает участника и наносит на рану выделения P. bicolor (Daly et al., 1992). Для сбора яда они отправляются на поиски лягушки на рассвете, ориентируясь на характерный звук. Благодаря медленному движению лягушки ее легко поймать (Daly et al., 1992; Den Brave et al., 2014). После поимки амфибию растягивают в форме буквы «Х» на скрещенных ветвях со связанными задними и передними лапами (Daly et al., 1992; Den Brave et al., 2014); однако некоторые жители извлекают яд, просто взяв животное в руки (рис. 5А). Сборщик дразнит амфибию тычками, что приводит к выделению кожного секрета. Выделяющийся секрет собирается путем осторожного соскабливания кожи лягушки деревянным стержнем и затем хранится на палочке для последующего использования (рис. 5A,B). После сбора выделений лягушку отпускают, и в целом не принято причинять ей вред, удалять чрезмерное количество яда или даже оставлять животное в ловушке на длительное время (Den Brave et al., 2014). Коренные собиратели считают, что причинение вреда может раздражать духов животных (Daly et al., 1992; Lima, 2005; Den Brave et al., 2014; Hesselink, 2018b; Majić et al., 2021).
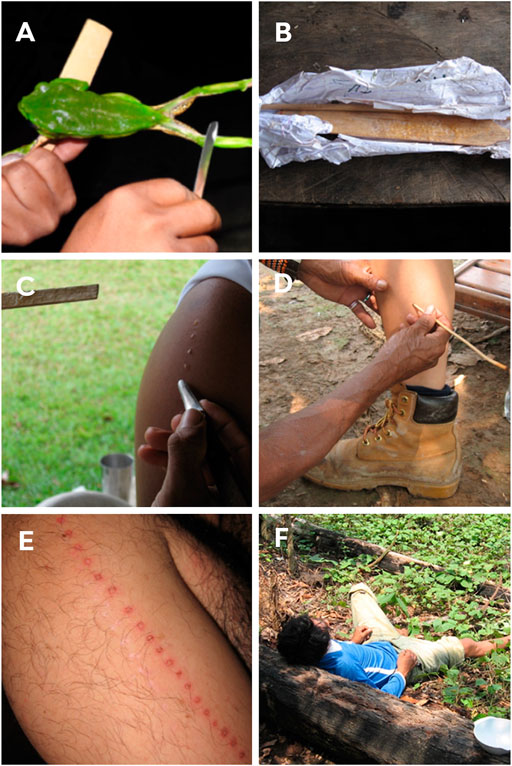
Ожоги делаются в виде точек с помощью тонкой лозы (так называемой «титики») (рис. 5D). Количество точек и место нанесения различны и связаны с тем, какая часть тела требуется для выполнения действий, выполняемых мужчинами и женщинами, а также учитывают, сколько раз участник подвергался ритуалу камбо (Lima, 1994; Hesselink, 2018b). Катукинцы наносят выделение на руки (дельтовидные) (рис. 5C,E) и грудь мужчин. У женщин она наносится на заднюю часть ног (икры) (рис. 5D), что объясняется необходимостью укрепить часть тела, наиболее привычную для их социальной роли, и участник может получить более ста «точек» одновременно (Bernarde and Santos, 2009). В прошлом это число могло быть втрое больше. Kaxinawá используют выделения в меньшем количестве на ногах или руках; они наносят от 2 до 10 «точек», в то время как Yawanawá — от 50 до 60 «точек» (Lima, 1994; Lima, 2000; Lima, 2005; Lima and Labate, 2008). Недавнее исследование, проведенное среди коренного населения Западной Амазонии (Рондония, Бразилия), показало, что женщины получают меньше «точек»: в среднем 8,2 и 10,3 «точек» при первом и втором применении, соответственно, тогда как мужчины — 8,6 и 11, соответственно.
Применение секреции обычно происходит на рассвете, и участникам рекомендуется выпить примерно 1-2 л свежего маниокового напитка или просто воды, а также поститься с вечера. При полном желудке участник обычно испытывает позыв к рвоте (Ribeiro, 2017). Сообщается, что реакция наступает через несколько минут после приема дозы. Реакция часто бывает сильной и включает тахикардию, потливость и сильную рвоту, но обычно проходит примерно через 60 мин, после чего наступает состояние апатии и сонливости (рис. 5F), которое может длиться от одной до нескольких минут, дней. Впоследствии, однако, участник сообщает о большей устойчивости и ясности мыслей (Daly et al., 1992; Hesselink, 2018c; Thompson et al., 2022).
Были проведены антропологические, биохимические и фармакологические исследования, однако до сих пор о ритуале камбо известно мало (Anastasi and Erspamer-Falconieri, 1970; Carneiro, 1970; Lima and Labate, 2008; Hesselink, 2018a; Hesselink and Winkelman, 2019). Первые этнографические наблюдения за ритуалом камбо у коренного населения верхней и средней части Джуруа были проведены французским миссионером Константином Тастевином в 1925 году (Tastevin, 1925). Предполагается, что примитивно-мифологическое использование секрета связано с шаманом из племени касинауа в Бразилии, который, видя множество тяжелобольных туземцев и не получая исцеления от ритуалов с аяхуаской, получил послание уйти в лес. Там он нашел божество, держащее в руках зеленую лягушку, и ему было открыто, как извлечь из лягушки секрецию и как ее применить (Hesselink, 2018c). Ритуал был успешно проведен и вошел в обиход касинауа и других племен (Lagrou, 1991; Lima and Labate, 2008; Venâncio and Melo-Sampaio, 2010; Hesselink, 2018c). Начиная с XIX века сосуществование коренных жителей и каучуконосов в Амазонии, в частности в долине реки Журуа, привело к богатому обмену знаниями и практиками и, в частности, к внедрению ритуала камбо речниками и каучуконосами (Balée, 2004). Франсиско Гомешу Мунису приписывают распространение ритуала камбо среди обработчиков каучука на притоке реки Журуа в Акко и первое применение секреции в городских центрах (Lima and Labate, 2008; Hesselink and Winkelman, 2019).
В XXI веке эта практика распространилась на другие неошаманские контуры и в настоящее время широко распространена в терапевтических клиниках, особенно связанных с альтернативными методами лечения и бразильскими религиями аяхуаски, то есть среди сторонников Санто Дайме и Униао ду Вегетал (Lima, 2005; Lopes, 2005; Lima and Labate, 2007; Lima and Labate, 2008; Bernarde and Santos, 2009). Однако в ходе распространения в городских центрах ритуал камбо трансформировался в терапевтические подходы и неошаманский ритуал исцеления, и этот процесс был назван «шаманизацией камбо», однако о ритуальной связи между этими двумя практиками ничего не известно (Lima and Labate, 2007; Hesselink, 2018c; Schmidt et al., 2020; Majić et al., 2021).
Распространение ритуала камбо среди некоренного населения Америки и Европы (Carneiro, 1970; Leban et al., 2016; Pogorzelska and Łapiński, 2017; Aquila et al., 2017; Li et al., 2018) выдвинуло на первый план обсуждение вопросов, связанных с традиционными знаниями, в частности озабоченность ответственностью знатоков традиций за неправильное применение секреции нетрадиционным населением. Silva et al. (2019) предупреждают, что в регионах, удаленных от мест традиционного применения вакцины, ее могут применять специалисты, не имеющие того же опыта, что и традиционное население, которое ее применяет, что создает риск для здоровья. Среди некоренных жителей Запада ритуал камбо проводится в церемониальном контексте, отличном от того, что происходит среди коренного населения, с пением, музыкальными инструментами, сжиганием благовоний и молитвами, рапе(содержащий Nicotiana rustica) и сананга (глазные капли из Tabernaemontana undulata), оба традиционных лекарства, часто применяются вместе с ритуалом камбо (Thompson and Williams, 2022). Продажа и применение секреции может привести к серьезным рискам для здоровья пользователей. В 2004 г. Бразильское национальное агентство по надзору за здравоохранением приняло решение о приостановке рекламы этой терапевтической альтернативы, поскольку не существует научных доказательств, гарантирующих качество, безопасность и эффективность секреции при любом типе расстройства, дисбаланса или лечения любых острых и хронических заболеваний (ANVISA, 2004).
Помимо нетрадиционного использования, поднимаются также вопросы, связанные с биопиратством (Daly et al., 1992; Lima, 2014a; Den Brave et al., 2014; Pogorzelska and Łapiński, 2017; Hesselink, 2018c; Peleg Hasson et al., 2021). Высушенные выделения P. bicolor на деревянных палочках коммерчески доступны как «палочки камбо» на рынках и в Интернете. Хотя использование секреции, по-видимому, остается устойчивым, расширение ее использования в последние годы может представлять собой значительное воздействие на окружающую среду и потенциальное сокращение численности вида (Cunha, 2009; Martins, 2010; Ribeiro, 2017). В 2003 г. катукинцы под руководством касика Фернандо выступили с осуждением неправомерного использования секрета и обвинили фармацевтическую промышленность в том, что она запатентовала пептиды камбо, а также заявила о своих правах на медицинские знания о секрете. В 2004 году был создан альянс между Катукинами и правительством Бразилии, чтобы гарантировать, что прибыль, полученная от разработки секреции, пойдет на благо Бразилии. В результате возник полевой проект, целью которого было объединение усилий аборигенов и исследователей, в основном направленных на регулирование использования секреции неаборигенами, а также на валоризацию и экономическое использование традиционных знаний о применении секреции. Однако проект не был реализован из-за сложности вопросов, связанных с правом собственности и распределением выгод (Lima, 2005; Lima, 2014b). С 2002 по 2018 год было утверждено 11 международных патентов, основанных на традиционных знаниях и природных генетических ресурсах, и все они были выданы странам Северного полушария, что может отражать присвоение природных генетических ресурсов и традиционных знаний (Shiva, 2001; Feres, 2022).
4 Медицинские аспекты энвомации P. bicolor
Причины, связанные с использованием, структурой и организацией ритуалов, а также психологические эффекты секреции у западных пользователей немногочисленны и остаются неподтверждёнными (Lima and Labate, 2008; Majić et al., 2021). Мажич и др. (2021) провели интернет-опрос, изучая различные аспекты использования секреции, и отметили множество мотивов ее применения, включая общее оздоровление, детоксикацию и духовный рост. По мнению Бернарде и Сантоса (2009 г.), причины, побуждающие к использованию секреции коренным населением западной Амазонии (Рондония, Бразилия), в целом сводятся к улучшению качества жизни и решению проблем со здоровьем, таким как ревматизм и диабет. Высказывались предположения о пользе для психического и физического здоровья, включая лечение различных патологий, таких как зависимость, депрессия, хроническая боль, аутоиммунные расстройства, тиреоидит Хашимото, целиакия, посттравматическое стрессовое расстройство, диабет, инфекционные заболевания, рак, гипертония и другие состояния здоровья (Hesselink, 2018a; c; Silva et al., 2019; Majić et al., 2021; Thompson and Williams, 2022). Хотя палочки, содержащие выделения лягушек, до сих пор можно заказать через интернет-сайты без рецепта, польза и важные биологические эффекты не доказаны в литературе, и она до сих пор не признана официально лекарственным средством. Полезные эффекты не были научно проверены в рандомизированных контролируемых исследованиях, поэтому лечебный эффект может быть просто эффектом плацебо (Aquila et al., 2017).
Механизмы действия и эффекты выделений P. bicolor до конца не изучены, особенно из-за разнообразия токсинов, входящих в состав выделений. In vitro были выявлены цитотоксическое и противоопухолевое действие на различные клетки, антибактериальные и противопротозойные свойства, а также вещества, обладающие высоким сродством к µ-опиоидным рецепторам (Negri et al., 1992; Erspamer et al, 1993; Leite et al., 2005; Hesselink and Schatman, 2018), а также расширение и повышение проницаемости сосудов и гематоэнцефалического барьера (Erspamer et al., 1989; Erspamer et al., 1993; Daly et al., 2009; Van Zoggel et al., 2012; Li et al., 2018). В целом выделения лягушек, могут вызвать серьезные побочные эффекты, вплоть до летального исхода. Кратковременные эффекты проявляются в течение нескольких минут и включают в себя желудочно-кишечные проблемы, характеризующиеся в основном сильной рвотой и диареей, мочеиспускание, потливость и тахикардию (Erspamer et al., 1989; Erspamer et al., 1993). Хотя некоторые эффекты считаются обязательными симптомами, поскольку они связаны с очищением организма от вредных влияний или токсинов (Hesselink, 2018d), эти эффекты, как правило, зависят от дозы (Daly et al., 1992; Li et al., 2018). Яркими примерами поздних эффектов секреции являются увеличение физической силы, выносливости и способности справляться со стрессовыми ситуациями. Другими иногда встречающимися эффектами являются галлюцинации, сонливость, головокружение, эйфория и седация (Erspamer et al., 1989; Erspamer et al., 1993). Эти эффекты опосредованы действием пептидов, присутствующих в кожном секрете, таких как филлокинин, адренорегулин, совагин, филлоцерулин, филломедузин, дерморфины и дельторфины (Li et al., 2018; Byard, 2019). Некоторые исследователи пытались понять весь эффект секреции, но получили немногочисленные и неопределенные результаты (Thompson et al., 2022; Thompson and Williams, 2022). В целом большинство исследований посвящено пептидам, полученным из секрета, а также отдельным эффектам основных выделенных компонентов (табл. 2).
Наиболее распространенные причины заболеваний и смерти после ритуалов камбо связаны с депрессивным воздействием производных опиоидов на ЦНС и влиянием токсинов на сердечно-сосудистую систему. Могут повреждаться почки, поджелудочная железа и печень (Hemingway et al., 2009; Pogorzelska and Łapiński, 2017). Имеется несколько сообщений об интоксикации как секретом P. bicolor, так и аяхуаской, которые привели к госпитализации (Morais et al., 2018; Campodónico et al., 2019). Однако есть и сообщения о госпитализации в связи с интоксикацией только секретом, которая сопровождалась тяжелыми побочными эффектами (Leban et al., 2016; Hesselink, 2018d; Campodónico et al., 2019; De la Vega et al., 2020; Sacco et al., 2022). В исследовании с участием 127 человек, употреблявших секрет, Thompson et al. (2022) наблюдали нетяжелые физические реакции, такие как отек лица, диафорез, расстройство кишечника и обморок, причем увеличение дозы вызывало усиление отека лица и потоотделения. Хотя другие исследования показали, что ритуал камбо может повышать риски повреждений от гипонатриемии, асфиксии от рвоты, а также травм от обморока и смерти (Menocchi, 2008; Labate and Lima, 2007; Aquila et al., 2017; Sacco et al., 2022), из-за высокой концентрации биоактивных пептидов и потенцирования определенных рецепторов эффекты могут быть ошибочно интерпретированы как интоксикация или массивная аллергическая реакция (Hesselink, 2018d).
5 Перспективы исследования
Являясь богатым источником пептидов, P. bicolor продолжает привлекать исследователей в области медицины и биотехнологий благодаря своему огромному химическому разнообразию (пока еще не до конца изученному), а также всем своим фармакологическим возможностям. Что касается этого потенциала, то необходимы исследования, направленные на дальнейшее выяснение механизмов действия некоторых пептидов, особенно опиоидов, которые могут найти применение в лечении хронических заболеваний. Параллельно следует продолжить изучение результатов исследований, свидетельствующих об антибиотическом, антинеопластическом, противовирусном, иммуномодулирующем и противопаразитарном потенциале. Необходимость дальнейших исследований подтверждается такими присущими этим пептидам характеристиками, как низкая молекулярная масса, высокая активность, низкая цитотоксичность, высокая водорастворимость и быстрая абсорбция. Таким образом, можно ожидать, что в будущем химия выделений P. bicolor продвинется к полезным и безопасным методам лечения, где традиционные знания вновь помогут будущим поколениям.
6 Заключительные замечания
Вид P. bicolor распространен в биоме Амазонки, где используется коренным населением в народном и традиционном здравоохранении. В настоящее время его применение распространилось и на другие группы населения, по-прежнему преследуя первоначальные цели профилактики и лечения ряда заболеваний и традиционных синдромов, а также для использования в некоторых религиозных культах. Расширение масштабов применения препарата вызывает опасения по поводу биопиратства: имеются сообщения о его использовании в европейских странах, России и США. Чрезмерное использование или использование лицами, имеющими ранее проблемы со здоровьем, может привести к тяжелым формам энвеноматоза и летальным исходам. Протоколов лечения таких случаев энвомации не существует, и в немногочисленных сообщениях, встречающихся в литературе, применяется поддерживающая терапия. Богатый состав биологически активных пептидов секрета, обладающих различной биологической активностью, обусловливает как признаки и симптомы их популярного применения, так и случаи энвеномании. Для лучшего понимания химической природы этих пептидов и их биологического действия на основе данных in vitro и ограниченных данных in vivo был проведен ряд исследований. С точки зрения биотехнологии интерес представляют антибиотические, антинеопластические, противовирусные, иммуномодулирующие и противопаразитарные потенциальные активности. (Bahar and Ren, 2013, Mor et al., 1994c, Vanhoye et al., 2004).
Авторский вклад
IK, MS, MP, PB, HK и WM разработали основную идею данной работы. Все авторы разработали и написали большинство тем данного обзора. TN, AB и MJ разработали рисунки данной обзорной статьи. Все авторы исправили рукопись и внесли важный вклад в развитие данной работы. Все авторы внесли свой вклад в подготовку статьи и одобрили ее окончательный вариант.
Финансирование
Мы благодарим Национальный совет по научным и техническим исследованиям (CNPq) за стипендии, предоставленные MP (№ 307184/2020-0), WM (№ 309207/2020-7), HK (№ 305942/2020-4), IK (№ 309473/2019-5) и PB (№ 311509/2020-7). WM признает финансирование со стороны Fundação de Amparo à Pesquisa do Estado do Amazonas (проекты Pro-Estado и Posgrad). MP (координатор проекта Snakebite Roraima) выражает благодарность за поддержку со стороны Hamish Ogston Foundation-Global Snakebite Initiative. HK была оказана поддержка по линии CT&I Priority Areas Call FAPEAM 010/2021 и POSGRAD 2020-2021. JS выражает признательность за финансирование со стороны Fundação de Amparo à Pesquisa do Estado do Amazonas (Science in Public Management Program-PCGP/FAPEAM 011/2021). Данное исследование было частично поддержано бразильской организацией CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior).
Конфликты интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все утверждения, высказанные в данной статье, принадлежат исключительно авторам и не обязательно отражают точку зрения их аффилированных организаций, а также издателя, редакторов и рецензентов. Любой продукт, который может быть оценен в данной статье, или утверждение, которое может быть сделано его производителем, не гарантируется и не поддерживается издателем.
Источники
Источники
Agüero-González, D. L., Pané-Vila, A., Gil, V., and Castro, P. (2019). Rituales de purificación con una rana amazónica (Kambô): Una peligrosa realidad en la Europa del siglo XXI. Emergencias 31 (5).
Alamos, M. F., Walker, R. H., and Miranda, M. (2020). Life-threatening risk of using Kambó in alternative medicine. Neurol. Clin. Pract. 10 (4), 1212 e35. doi:10.1212/CPJ.0000000000000749
PubMed Abstract | CrossRef Full Text | Google Scholar
Altig, R., and McDiarmid, R. W. (1999). “Tadpoles: The biology of Anuran Larvae,” in Body plan: Development and morphology. Editors R. W. McDiarmid, and R. Altig (The University of Chicago Press), 24–51.
Amiche, M., Seon, A. A., Wroblewski, H., and Nicolas, P. (2000). Isolation of dermatoxin from frog skin, an antibacterial peptide encoded by a novel member of the dermaseptin genes family. Eur. J. Biochem. 267 (14), 4583–4592. doi:10.1046/j.1432-1327.2000.01514.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Anastasi, A., Bertaccini, G., Cei, J. M., De Caro, G., Erspamer, V., and Impicciatore, M. (1969). Structure and pharmacological actions of phyllocaerulein, a caerulein-like nonapeptide: Its occurrence in extracts of the skin of Phyllomedusa sauvagei and related Phyllomedusa species. Br. J. Pharmacol. 37 (1), 198–206. doi:10.1111/j.1476-5381.1969.tb09538.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Anastasi, A., Bertaccini, G., and Erspamer, V. (1966). Pharmacological data on phyllokinin (bradykinyl‐ isoleucyl‐ tyrosine o‐ sulphate) and bradykinyl‐ isoleucyl‐ tyrosine. Br. J. Pharmacol. Chemother. 27 (3), 479–485. doi:10.1111/j.1476-5381.1966.tb01859.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Anastasi, A., and Falconieri Erspamer, G. (1970). Occurrence of phyllomedusin, a physalaemin-like decapeptide, in the skin of Phyllomedusa bicolor. Experientia 26 (8), 866–867.
PubMed Abstract | CrossRef Full Text | Google Scholar
ANVISA (2004). Resolução-RE Nº8, de 29 de Abril de 2004. Agência Nacional de Vigilância Sanitária. Diário Of. União, Brasília, DF, Seção 1 Nº 82, 94, 30 abr.
Aquila, I., Gratteri, S., Sacco, M. A., Fineschi, V., Magi, S., Castaldo, P., et al. (2017). The biological effects of kambo: Is there a relationship between its administration and sudden death? J. Forensic Sci. 63 (3), 965–968. doi:10.1007/BF02114227
PubMed Abstract | CrossRef Full Text | Google Scholar
Bahar, A. A., and Ren, D. (2013). Antimicrobial peptides. Pharmaceuticals 6 (12), 1543–1575. doi:10.3390/ph6121543
PubMed Abstract | CrossRef Full Text | Google Scholar
Balée, W. (2004). Enciclopédia da floresta: o alto Juruá: Práticas e conhecimentos das populações. Mana 10 (2), 419–423. doi:10.1590/S0104-93132004000200009
CrossRef Full Text | Google Scholar
Barra, D., and Simmaco, M. (1995). Amphibian skin: A promising resource for antimicrobial peptides. Trends Biotechnol. 13 (6), 205–209. doi:10.1016/S0167-7799(00)88947-7
PubMed Abstract | CrossRef Full Text | Google Scholar
Basir, Y. J., Knoop, F. C., Dulka, J., and Conlon, J. M. (2000). Multiple antimicrobial peptides and peptides related to bradykinin and neuromedin N isolated from skin secretions of the pickerel frog, Rana palustris. Biochimica Biophysica Acta (BBA)-Protein Struct. Mol. Enzym. 1543 (1), 95–105. doi:10.1016/S0167-4838(00)00191-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Bernarde, P. S. (2012). Anfíbios e répteis: Introdução ao estudo da herpetofauna brasileira. Curitiba: Anolis Books.
Bernarde, P. S., and Santos, R. A. (2009). Utilização medicinal da secreção (“vacina-do-sapo”) do anfíbio kambô (Phyllomedusa bicolor)(Anura: Hylidae) por população não-indígena em Espigão do Oeste, Rondônia, Brasil. Biotemas 22 (3), 213–220. doi:10.5007/2175-7925.2009v22n3p213
CrossRef Full Text | Google Scholar
Bertoluci, J. (2002). Pedal luring in the leaf-frog Phyllomedusa burmeisteri (Anura, hylidae, phyllomedusinae). Phyllomedusa J. Herpetology 1 (2), 93–95. doi:10.11606/issn.2316-9079.v1i2p93-95
CrossRef Full Text | Google Scholar
Bevins, C. L., and Zasloff, M. (1990). Peptides from frog skin. Annu. Rev. Biochem. 59 (1), 395–414. doi:10.1146/annurev.bi.59.070190.002143
PubMed Abstract | CrossRef Full Text | Google Scholar
Blaylock, L. A., Ruibal, R., and Platt-Aloia, K. (1976). Skin structure and wiping behavior of Phyllomedusinae frogs. Copeia 1976 (2), 283–295. doi:10.2307/1443948
CrossRef Full Text | Google Scholar
Boddaert, P. (1772). Over den Twee-koleurigen Kikvirsch. De Rana Bicolore 41.
Broccardo, M., Erspamer, V., Falconieri, G., Improta, G., Linari, G., Melchiorri, P., et al. (1981). Pharmacological data on dermorphins, a new class of potent opioid peptides from amphibian skin. Br. J. Pharmacol. 73 (3), 625–631. doi:10.1111/j.1476-5381.1981.tb16797.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Buchacher, C. O. (1993). Field studies on the small Surinam toad, Pipa arrabali, near Manaus, Brazil. Amphibia-Reptilia 14 (1), 59–69. doi:10.1163/156853893×00192
CrossRef Full Text | Google Scholar
Byard, R. W. (2019). Is voluntary envenomation from the kambô ritual therapeutic or toxic? Forensic Sci. Med. Pathology 16 (2), 205–206. doi:10.1007/s12024-019-00192-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Calderon, L. A., Silva, A. A. E., Ciancaglini, P., and Guerino, S. R. (2011). Antimicrobial peptides from Phyllomedusa frogs: From biomolecular diversity to potential nanotechnologic medical applications. Amino Acids 40, 29–49. doi:10.1007/s00726-010-0622-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Campião, K. M., Morais, D. H., Dias, O. T., Aguiar, A., Toledo, G. M., Tavares, L. E. R., et al. (2014). Checklist of helminth parasites of amphibians from South America. Zootaxa 3843 (1), 1–93. doi:10.11646/zootaxa.3843.1.1
PubMed Abstract | CrossRef Full Text | Google Scholar
Campodónico, J., Aedo, P., Montané, M. I., Rojas, A., Aveiga, A., Silva, L., et al. (2019). Severe hyponatremia secondary to Phyllomedusa bicolor (Kambó frog) poisoning. Report of one case. Rev. Médica Chile 147, 935–939.
Cao, W., Zhou, Y., Ma, Y., Luo, Q., and Wei, D. (2005). Expression and purification of antimicrobial peptide adenoregulin with C-amidated terminus in Escherichia coli. Protein Expr. Purif. 40 (2), 404–410. doi:10.1016/j.pep.2004.12.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Caramaschi, U., and Cruz, C. A. G. (2002). Phyllomedusa: Posição taxonômica, hábitos e biologia (Amphibia, Anura, hylidae). Phyllomedusa 1, 5–10. doi:10.11606/issn.2316-9079.v1i1p05-10
CrossRef Full Text | Google Scholar
Carneiro, R. L. (1970). Hunting and hunting magic among the amahuaca of the Peruvian Montana. Ethnology 9 (4), 331–341. doi:10.2307/3773039
CrossRef Full Text | Google Scholar
Castanho, L. M., and de Luca, I. M. S. (2001). Moulting behavior in leaf-frogs of the genus Phyllomedusa (Anura: Hylidae). Zoologischer Anzeiger-A J. Comp. Zoology 240 (1), 3–6. doi:10.1078/0044-5231-00001
CrossRef Full Text | Google Scholar
Castroviejo-Fisher, S., Köhler, J., De la Riva, I., and Padial, J. M. (2017). A new morphologically cryptic species of Phyllomedusa (Anura: Phyllomedusidae) from Amazonian forests of northern Peru revealed by DNA sequences. Zootaxa 4269, 245–264. doi:10.11646/ZOOTAXA.4269.2.4
PubMed Abstract | CrossRef Full Text | Google Scholar
Chai, N. (2015). Endoscopy in Amphibians. Veterinary Clin. N. Am Exot. Animal Pract. 18 (3), 479–491. doi:10.1016/j.cvex.2015.04.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Charpentier, S., Amiche, M., Mester, J., Vouille, V., Le Caer, J. P., Nicolas, P., et al. (1998). Structure, synthesis, and molecular cloning of dermaseptins B, a family of skin peptide antibiotics. J. Biol. Chem. 273 (24), 14690–14697. doi:10.1074/jbc.273.24.14690
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, F. Y., Lee, M. T., and Huang, H. W. (2002). Sigmoidal concentration dependence of antimicrobial peptide activities: A case study on alamethicin. Biophysical J. 82 (2), 908–914. doi:10.1016/S0006-3495(02)75452-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, Z., Xi, X., Lu, Y., Hu, H., Dong, Z., Ma, C., et al. (2021). In vitro activities of a novel antimicrobial peptide isolated from Phyllomedusa tomopterna. Microb. Pathog. 153, 104795. doi:10.1016/j.micpath.2021.104795
PubMed Abstract | CrossRef Full Text | Google Scholar
Clarke, B. T. (1997). The natural history of Amphibian skin secretions, their normal functioning and potential medical applications. Biol. Rev. Camb. Philosophical Soc. 72 (3), 365–379. doi:10.1017/s0006323197005045
PubMed Abstract | CrossRef Full Text | Google Scholar
Cline, M. A., Cofield, S. A., and Tachibana, T. (2010). Central litorin injection is associated with primary anorexigenic effects that coincide with activation of the magnocellular division of the paraventricular nucleus. Neuropeptides 44 (3), 247–252. doi:10.1016/j.npep.2009.12.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Conceição, K., Konno, K., Richardson, M., Antoniazzi, M. M., Jared, C., Daffre, S., et al. (2006). Isolation and biochemical characterization of peptides presenting antimicrobial activity from the skin of Phyllomedusa hypochondrialis. Peptides 27 (12), 3092–3099. doi:10.1016/j.peptides.2006.08.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Conibear, A. C. (2020). Deciphering protein post-translational modifications using chemical biology tools. Nat. Rev. Chem. 4, 674–695. doi:10.1038/s41570-020-00223-8
CrossRef Full Text | Google Scholar
Cunha, M. C. (2009). “Cultura e cultura: Conhecimentos tradicionais e direitos intelectuais,” in Cultura com aspas (São Paulo: Cosac Naify), 311–373.
Daly, J. W., Caceres, J., Moni, R. W., Gusovsky, F., Moos, M., Seamon, K. B., et al. (1992). Frog secretions and hunting magic in the upper Amazon: Identification of a peptide that interacts with an adenosine receptor. Proc. Natl. Acad. Sci. 89 (22), 10960–10963. doi:10.1073/pnas.89.22.10960
PubMed Abstract | CrossRef Full Text | Google Scholar
Daly, J. W. (1995). The chemistry of poisons in amphibian skin. Proc. Natl. Acad. Sci. 92 (1), 9–13. doi:10.1073/pnas.92.1.9
PubMed Abstract | CrossRef Full Text | Google Scholar
Daly, J. W., Ware, N., Saporito, R. A., Spande, T. F., and Garraffo, H. M. (2009). N-Methyldecahydroquinolines: An unexpected class of alkaloids from amazonian poison frogs (Dendrobatidae). J. Nat. Prod. 72 (6), 1110–1114. doi:10.1021/np900094v
PubMed Abstract | CrossRef Full Text | Google Scholar
de la Vega, M., Maldonado, G., and Kraus, A. (2020). Dermatomyositis induced by the secretion of Phyllomedusa bicolor or Kambô frog — a case report. Toxicon 184, 57–61. doi:10.1016/j.toxicon.2020.05.018
PubMed Abstract | CrossRef Full Text | Google Scholar
Delfino, G. (1991). in Ultrastructural aspects of venom secretion in anuran cutaneous glands”Handbook of natural toxins. Editors A. T. Tu, and M. Dekker (New York, NY, USA: Marcel Dekker), 777–802.
Delfino, G., Brizzi, R., Alvarez, B. B., and Kracke-Berndorff, R. (1998). Serous cutaneous glands in Phyllomedusa hypochondrialis (Anura:hylidae): Secretory patterns during ontogenesis. Tissue Cell 30 (1), 30–40. doi:10.1016/S0040-8166(98)80004-9
PubMed Abstract | CrossRef Full Text | Google Scholar
den Brave, P. S., Bruins, E., and Bronkhorst, M. W. (2014). Phyllomedusa bicolor skin secretion and the Kambô ritual. J. Venom. Animals Toxins Incl. Trop. Dis. 20, 1–3. doi:10.1186/1678-9199-20-40
CrossRef Full Text | Google Scholar
Doyle, J., Llewellyn, L. E., Brinkworth, C. S., Bowie, J. H., Wegener, K. L., Rozek, T., et al. (2002). Amphibian peptides that inhibit neuronal nitric oxide synthase: The isolation of lesueurin from the skin secretion of the Australian stony creek frog Litoria lesueuri. Eur. J. Biochem. 269 (1), 100–109. doi:10.1046/j.0014-2956.2002.02630.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Duellman, W. E., Marion, A. B., and Hedges, S. B. (2016). Phylogenetics, classification, and biogeography of the treefrogs (Amphibia: Anura: Arboranae). Zootaxa 4104 (1), 1–109. doi:10.11646/ZOOTAXA.4104.1.1
PubMed Abstract | CrossRef Full Text | Google Scholar
Eckart, K., Radulovic, J., Radulovic, M., Jahn, O., Blank, T., Stiedl, O., et al. (1999). Actions of CRF and its analogs. Curr. Med. Chem. 6 (11), 1035–1053.
PubMed Abstract | CrossRef Full Text | Google Scholar
Erspamer, V., Falconieri-Erspamer, G., Severini, C., Potenza, R. L., Barra, D., Mignogna, G., et al. (1993). Pharmacological studies of “sapo” from the frog Phyllomedusa bicolor skin: A drug used by the Peruvian matses Indians in shamanic hunting practices. Toxicon 31, 1099–1111.
PubMed Abstract | CrossRef Full Text | Google Scholar
Erspamer, V., Melchiorri, P., Broccardo, M., Erspamer, G. F., Falaschi, P., Improta, G., et al. (1981). The brain-gut-skin triangle: New peptides. Peptides 2 (2), 7–16. doi:10.1016/0196-9781(81)90003-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Erspamer, V., Melchiorri, P., Falconieri-Erspamer, G., Montecucchi, P. C., and de Castiglione, R. (1985). Phyllomedusa skin: A huge factory and store-house of a variety of active peptides. Peptides 6 (3), 7–12. doi:10.1016/0196-9781(85)90343-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Erspamer, V., Melchiorri, P., Falconieri-Erspamer, G., Negri, L., Corsi, R., Severini, C., et al. (1989). Deltorphins: A family of naturally occurring peptides with high affinity and selectivity for delta opioid binding sites. Proc. Natl. Acad. Sci. 86 (13), 5188–5192. doi:10.1073/pnas.86.13.5188
PubMed Abstract | CrossRef Full Text | Google Scholar
Erspamer-Falconieri, G., Anastasi, A., and Cei, J. M. (1970). Pharmacological observations on phyllomedusin. J. Pharm. Pharmacol. 22 (6), 466–467. doi:10.1111/j.2042-7158.1970.tb08567.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Esakov, A. I., Ashmarin, I. P., Serova, O. N., Obukhova, M. F., Storozheva, Z. I., Golubovich, V. P., et al. (1990). Litorin and litorin-albumin conjugate as effective regulators of body temperature in rats. Biomed. Sci. 1 (6), 610–612.
PubMed Abstract | Google Scholar
Faivovich, J., Haddad, C. F. B., Baêta, D., Jungfer, K. H., Álvarez, G. R. F., Brandão, R. A., et al. (2010). The phylogenetic relationships of the charismatic poster frogs, Phyllomedusinae (Anura, Hylidae). Cladistics 26, 227–261. doi:10.1111/j.1096-0031.2009.00287.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Faivovich, J., Haddad, C. F. B., Garcia, P. C. A., Frost, D. R., Campbell, J. A., and Wheeler, W. C. (2005). Systematic review of the frog family hylidae, with special reference to hylinae: Phylogenetic analysis and taxonomic revision. Bull. Am. Mus. Nat. Hist. 294, 1–240. doi:10.5531/sd.sp.12
CrossRef Full Text | Google Scholar
Feder, R., Dagan, A., and Mor, A. (2000). Structure-activity relationship study of antimicrobial dermaseptin S4 showing the consequences of peptide oligomerization on selective cytotoxicity. J. Biol. Chem. 275 (6), 4230–4238.
PubMed Abstract | CrossRef Full Text | Google Scholar
Feres, M. V. C. (2022). Biodiversity, traditional knowledge and patent rights: The case study of Phyllomedusa bicolor. Rev. Direito GV 18 (1), e2205. doi:10.1590/2317-6172202205
CrossRef Full Text | Google Scholar
Fiori, A., Cardelli, P., Negri, L., Savi, M. R., Strom, R., and Erspamer, V. (1997). Deltorphin transport across the blood–brain barrier. Proc. Natl. Acad. Sci. 94 (17), 9469–9474. doi:10.1073/pnas.94.17.9469
PubMed Abstract | CrossRef Full Text | Google Scholar
Fleury, Y., Vouille, V., Beven, L., Amiche, M., Wróblewski, H., Delfour, A., et al. (1998). Synthesis, antimicrobial activity and gene structure of a novel member of the dermaseptin B family. Biochimica Biophysica Acta (BBA)-Gene Struct. Expr. 1396 (2), 228–236. doi:10.1016/S0167-4781(97)00194-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Frost, D. R. (2021). Amphibian species of the world: An online reference. Version6.1. Disponível em. New York, NY, USA: American Museum of Natural History. Acessed https://amphibiansoftheworld.amnh.org/index.php April 11, 2022.
Gehara, M., Crawford, A. J., Orrico, V. G., Rodriguez, A., Loetters, S., Fouquet, A., et al. (2014). High levels of diversity uncovered in a widespread nominal taxon: Continental phylogeography of the neotropical tree frog Dendropsophus minutus. PloS One 9 (9), e103958. doi:10.1371/journal.pone.0103958
PubMed Abstract | CrossRef Full Text | Google Scholar
Gomez-Mestre, I., Wiens, J. J., and Warkentin, K. M. (2008). Evolution of adaptive plasticity: Risk-sensitive hatching in neotropical leafbreeding treefrogs. Ecol. Monogr. 78, 205–224. doi:10.1890/07-0529.1
CrossRef Full Text | Google Scholar
Góngora-Benítez, M., Tulla-Puche, J., and Albericio, F. (2014). Multifaceted roles of disulfide bonds. Peptides as therapeutics. Chem. Rev. 114, 901–926. doi:10.1021/cr400031z
PubMed Abstract | CrossRef Full Text | Google Scholar
Gonzaga, E. S. R., Chamorro, M., Ganti, L., and Schneider, R. (2020). Kambo frog poison as a cause of esophageal rupture. Cureus 12 (9), e10677. doi:10.7759/cureus.10677
PubMed Abstract | CrossRef Full Text | Google Scholar
Govender, T., Dawood, A., Esterhuyse, A. J., and Katerere, D. R. (2012). Antimicrobial properties of the skin secretions of frogs. South Afr. J. Sci. 108 (5), 1. doi:10.10520/EJC120717
CrossRef Full Text | Google Scholar
Grandt, D., Schimiczek, M., Struk, K., Shively, J., Eysselein, V. E., Goebell, H., et al. (1994). Characterization of two forms of peptide YY, PYY (1–36) and PYY (3–36), in the rabbit. Peptides 15 (5), 815–820. doi:10.1016/0196-9781(94)90035-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Haddad, V., and Martins, I. A. (2020). Kambô: An amazonian enigma. J. Venom Res. 10, 13.
PubMed Abstract | Google Scholar
Hantak, M. M., Grant, T., Reinsch, S., Mcginnity, D., Loring, M., Toyooka, N., et al. (2013). Dietary alkaloid sequestration in a poison frog: An experimental test of alkaloid uptake in Melanophryniscus stelzneri (bufonidae). J. Chem. Ecol. 39 (11), 1400–1406. doi:10.1007/s10886-013-0361-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Hay, D. L., Garelja, M. L., Poyner, D. R., and Walker, C. S. (2017). Update on the pharmacology of calcitonin/CGRP family of peptides: IUPHAR review 25. Br. J. Pharmacol. 175 (1), 3–17. doi:10.1111/bph.14075
PubMed Abstract | CrossRef Full Text | Google Scholar
Heinrichs, S. C., and Taché, Y. (2001). Therapeutic potential of CRF receptor antagonists: A gut-brain perspective. Expert Opin. Investig. Drugs 10 (4), 647–659. doi:10.1517/13543784.10.4.647
PubMed Abstract | CrossRef Full Text | Google Scholar
Hemingway, V., Brunner, J., Speare, R., Berger, L., Heatwole, H., and Wilkinson, J. W. (2009). “Amphibian decline: Disease, parasites, maladies, and pollution,” in Amphibian Biology.
Hesselink, J. M. K. (2018a). Kambô: A shamanistic ritual arriving in the West: Description, risks and perception by the users. Int. J. Psychol. Psychoanal. 4 (2), 034. doi:10.23937/2572-4037.1510034
CrossRef Full Text | Google Scholar
Hesselink, J. M. K. (2018b). Kambo: A ritualistic healing substance from an amazonian frog and a source of new treatments. Open J. Pain Med. 2 (1), 4–006. doi:10.17352/ojpm.000007
CrossRef Full Text | Google Scholar
Hesselink, J. M. K. (2018c). Kambô: A shamanic medicine-personal testimonies. JOJ Case Stud. 8 (3), 555739. doi:10.19080/JOJCS.2018.08.555739
CrossRef Full Text | Google Scholar
Hesselink, J. M. K. (2018d). Kambo and its multitude of biological effects: Adverse events or pharmacological effects. Int. Archives Clin. Pharmacol. 4 (1), 17. doi:10.23937/2572-3987.1510017
CrossRef Full Text | Google Scholar
Hesselink, J. M. K., and Sacerdote, P. (2019). Frogs as a rich source for new treatment principles: A review of bio-active peptides in phyllomedusae (Phyllomedusa bicolor) and bombinae (Bombina variegate). SL Pharmacol. Toxicol. 2 (1), 116.
Hesselink, J. M. K., and Schatman, M. E. (2018). Rediscovery of old drugs: The forgotten case of dermorphin for postoperative pain and palliation. J. Pain Res. 11, 2991–2995. doi:10.2147/JPR.S186082
PubMed Abstract | CrossRef Full Text | Google Scholar
Hesselink, J. M. K., and Winkelman, M. (2019). Vaccination with Kambô against bad influences: Processes of symbolic healing and ecotherapy. J. Transpers. Psychol. 51 (1), 28–48.
Iakp (2022). International association of kambo practitioners. Available at: https://iakp.org/ (Acessed April 3, 2022).
IUCN (2021). The IUCN red list of threatened species. Version 2021-3 Accessed on: https://www.iucnredlist.org (Acessed April 11, 2022).
Jairam, R. (2019). Checklist of the amphibians and reptiles of the Lely Mountains, eastern Suriname. Amphibian Reptile Conservation 13 (2), e200.
Jared, C., Antoniazzi, M. M., Jordão, A. E., Silva, J. R. M., Greven, H., and Rodrigues, M. T. (2009). Parotoid macroglands in toad (Rhinella jimi): Their structure and functioning in passive defence. Toxicon 54 (3), 197–207. doi:10.1016/j.toxicon.2009.03.029
PubMed Abstract | CrossRef Full Text | Google Scholar
Jeckel, A. M., Saporito, R. A., and Grant, T. (2015). The relationship between poison frog chemical defenses and age, body size, and sex. Front. Zoology 12 (1), 1–9. doi:10.1186/s12983-015-0120-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, J. B., Johansson, Å., and Conlon, J. M. (2001). Anomalous rates of evolution of pancreatic polypeptide and peptide tyrosine-tyrosine (PYY) in a tetraploid frog, Xenopus laevis (Anura: Pipidae). Peptides 22 (3), 317–323. doi:10.1016/S0196-9781(01)00333-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Kimbrell, D. A., and Beutler, B. (2001). The evolution and genetics of innate immunity. Nat. Rev. Genet. 2 (4), 256–267. doi:10.1038/35066006
PubMed Abstract | CrossRef Full Text | Google Scholar
König, E., Clark, V. C., Shaw, C., and Bininda-Emonds, O. R. (2012). Molecular cloning of skin peptide precursor-encoding cDNAs from tibial gland secretion of the Giant Monkey Frog, Phyllomedusa bicolor (Hylidae, Anura). Peptides 38 (2), 371–376. doi:10.1016/j.peptides.2012.09.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Kückelhaus, S., Leite, J. R. S., Neves, M. P., Frota, K. S., Abdala, L. F., Muniz-Junqueira, M. I., et al. (2007). Toxicity evaluation to mice of phylloseptin-1, an antimicrobial peptide from the skin secretion of Phyllomedusa hypochondrialis (Amphibia). Int. J. Peptide Res. Ther. 13 (3), 423–429. doi:10.1007/s10989-006-9060-1
CrossRef Full Text | Google Scholar
Kückelhaus, S. A., Leite, J. R. S., Muniz-Junqueira, M. I., Sampaio, R. N., Bloch, C., and Tosta, C. E. (2009). Antiplasmodial and antileishmanial activities of phylloseptin-1, an antimicrobial peptide from the skin secretion of Phyllomedusa azurea (Amphibia). Exp. Parasitol. 123 (1), 11–16. doi:10.1016/j.exppara.2009.05.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Kumachev, A., Zipursky, J. S., Weinerman, A. S., and Thompson, M. (2018). Poisoning from the kambô ritual. Can. J. Emerg. Med. 20 (6), 962–964. doi:10.1017/cem.2018.58
CrossRef Full Text | Google Scholar
Labate, B. C., and Lima, E. C. (2014). Medical drug or shamanic power plant: The uses of kambô in Brazil. Ponto Urbe 15, 1–20. doi:10.4000/pontourbe.2384
CrossRef Full Text | Google Scholar
Lacombe, C., Cifuentes-Diaz, C., Dunia, I., Auber-Thomay, M., Nicolas, P., and Amiche, M. (2000). Peptide secretion in the cutaneous glands of South American tree frog Phyllomedusa bicolor: An ultrastructural study. Eur. J. Cell Biol. 79 (9), 631–641. doi:10.1078/0171-9335-00085
PubMed Abstract | CrossRef Full Text | Google Scholar
Ladram, A., and Nicolas, P. (2016). Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. 21, 1341–1371. doi:10.2741/4461
PubMed Abstract | CrossRef Full Text | Google Scholar
Lagrou, E. M. (1991). Uma etnografia da cultura Kaxinawá entre a cobra e o inca. Federal University of Santa Catarina. [Dissertation] [Florianópolis (SC)].
Larsen, E. H., and Ramløv, H. (2013). Role of cutaneous surface fluid in frog osmoregulation. Comp. Biochem. Physiology Part A Mol. Integr. Physiology 165 (3), 365–370. doi:10.1016/j.cbpa.2013.04.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Lazarus, L. H., Bryant, S. D., Cooper, P. S., and Salvadori, S. (1999). What peptides these deltorphins be. Prog. Neurobiol. 57 (4), 377–420. doi:10.1016/S0301-0082(98)00050-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Leban, V., Kozelj, G., and Brvar, M. (2016). The syndrome of inappropriate antidiuretic hormone secretion after giant leaf frog (Phyllomedusa bicolor) venom exposure. Toxicon 120, 107–109. doi:10.1016/j.toxicon.2016.07.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Leite, J. R. S., Silva, L. P., Rodrigues, M. I. S., Prates, M. V., Brand, G. D., Lacava, B. M., et al. (2005). Phylloseptins: A novel class of anti-bacterial and anti-protozoan peptides from the Phyllomedusa genus. Peptides 26 (4), 565–573. doi:10.1016/j.peptides.2004.11.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, K., Horng, H., Lynch, K., and Smollin, C. G. (2018). Prolonged toxicity from Kambo cleansing ritual. Clin. Toxicol. 56 (11), 1165–1166. doi:10.1080/15563650.2018.1457153
PubMed Abstract | CrossRef Full Text | Google Scholar
Lima, E. C., and Labate, B. C. (2007). Remédio da ciência» e» remédio da alma»: Os usos da secreção do kambô (Phyllomedusa bicolor) nas cidades. Campos — Rev. Antropol. 8 (1), 71–90. doi:10.5380/cam.v8i1.9553
CrossRef Full Text | Google Scholar
Lima, E. C., and Labate, B. C. (2008). A expansão urbana do kampo (Phyllomedusa bicolor): Notas etnográficas1. DroDrogas Cult. Novas Perspect., 315–344.
Lima, E. C. (1994). Katukina: Historia e organizacao social de um grupo pano do alto jurua. University of São Paulo. [Dissertation][São Paulo (SP)].
Lima, E. C. (2000). Detalhe do registro: Com os olhos da serpente: Homens, animais e espíritos nas concepções Katukina sobre a natureza. University of São Paulo. [Thesis][São Paulo (SP)].
Lima, E. C. (2005). Kampu, kampo, kambô: o uso do sapo-verde entre os katukina. Rev. do Patrimônio Histórico Artístico Nac. 32, 255–267.
Lima, E. C. (2008). “As novas formas do kampô: Elementos de uma sociologia da disseminação urbana dos saberes nativos,” in Pueblos indígenas, plantas y mercados–amazonía y gran chaco. V congreso CEISAL de Latinoamericanistas, zeta series in anthropology & sociology.
Lima, E. C. (2009). “Entre o mercado esotérico e os direitos de propriedade intelectual: o caso do kampô (Phyllomedusa bicolor),” in Dilemas do acesso à biodiversidade e aos conhecimentos tradicionais. Editors J. Kleba, and S. Kishi, (Belo Horizonte, BH: Direito. política e sociedade).
Lima, E. C. (2012). A gente é que sabe” ou sobre as coisas katukina (pano). Rev. Antropol. 55 (1), 139–169. doi:10.11606/2179-0892.ra.2012.46962
CrossRef Full Text | Google Scholar
Lima, E. C. (2014a). Um objeto ou uma técnica? O caso do kampô. Rev. Antropol. UFSCar 6 (2), 21–33. doi:10.52426/rau.v6i2.120
CrossRef Full Text | Google Scholar
Lima, E. C. (2014b). “A internacionalização do kampô (via ayahuasca): Difusão global e efeitos locais,” in Políticas culturais e povos indígenas. Editors M. C. Cunha, and P. N. Cesarino (São Paulo, SP: Cultura Acadêmica)), 91–112.
Lima, A. P., Magnusson, W. E., Menin, M., Erdtmann, L. K., Rodrigues, D. D. J., Keller, C., et al. (2012). Guia de sapos da Reserva Adolpho Ducke-Amazônia Central. São Paulo: Áttema Design Editorial.
Liu, Y., Shi, D., Wang, J., Chen, X., Zhou, M., Xi, X., et al. (2020). A novel amphibian antimicrobial peptide, phylloseptin-PV1, exhibits effective anti-staphylococcal activity without inducing either hepatic or renal toxicity in mice. Front. Microbiol 11, 565158. doi:10.3389/fmicb.2020.565158
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, H., Guo, X., Yi, T., Zhu, Y., Ren, X., Guo, R., et al. (2021). Frog skin derived peptides with potential protective effects on ultraviolet B–induced cutaneous photodamage. Front. Immunol. 12, 2127. doi:10.3389/fimmu.2021.613365
CrossRef Full Text | Google Scholar
Long, Q., Wang, L., Zhou, M., Wu, Y., and Chen, T. (2018). A novel peptide Phylloseptin‐PB u from Phyllomedusa burmeisteri possesses insulinotropic activity via potassium channel and GLP‐1 receptor signalling. J. Cell. Mol. Med. 22 (5), 2804–2814. doi:10.1111/jcmm.13573
PubMed Abstract | CrossRef Full Text | Google Scholar
Lopes, L. A. (2005). Kambô, a medicina da floresta (experiência narrativa). [Monography][São Paulo (SP)]. University of São Paulo.
Lovejoy, D. A., and Balment, R. J. (1999). Evolution and physiology of the corticotropin-releasing factor (CRF) family of neuropeptides in vertebrates. General Comp. Endocrinol. 115 (1), 1–22. doi:10.1006/gcen.1999.7298
PubMed Abstract | CrossRef Full Text | Google Scholar
Majić, T., Sauter, M., Bermpohl, F., and Schmidt, T. T. (2021). Connected to the spirit of the frog: An Internet-based survey on Kambô, the secretion of the Amazonian Giant Maki Frog (Phyllomedusa bicolor): Motivations for use, settings and subjective experiences. J. Psychopharmacol. Oxf. Engl. 35 (4), 421–436. doi:10.1177/0269881121991554
CrossRef Full Text | Google Scholar
Martins, H. M. (2010). “Os katukina e o kampô: Percursos de um «conhecimento tradicional» indígena,” in Conhecimentos tradicionais para o século XXI: Etnografias da intercientificidade. Editors P. E. Little, and D. F. Braília (Brasilia: IEB Press), 35–73.
Martins, M., and Oliveira, M. E. (1998). Natural history of snakes in forests of the Manaus region, Central Amazonia, Brazil. Herpetol. Nat. Hist. 6 (2), 78–150.
Mathison, R., and Solomos, D. (1985). Interaction of tachykinins with an adrenergic receptor in the rat urinary bladder. Eur. J. Pharmacol. 109 (3), 327–333. doi:10.1016/0014-2999(85)90392-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Menin, M., Waldez, F., and Lima, A. P. (2008). Temporal variation in the abundance and number of species of frogs in 10,000 ha of a forest in Central Amazonia, Brazil. S. Am. J. Herpetology 3, 682–781. doi:10.2994/1808-9798(2008)3[68:TVITAA]2.0.CO;2
CrossRef Full Text | Google Scholar
Menocchi, S. (2008). Investigada morte de homem que tomou “vacina do sapo”. Estadão (São Paulo). Available at: https://www.estadao.com.br/noticias/geral,investigada-morte-de-homem-que-tomou-vacina-do-sapo, 162933
Mignogna, G., Severini, C., Simmaco, M., Negri, L., Erspamer, G. F., Kreil, G., et al. (1992). Identification and characterization of two dermorphins from skin extracts of the Amazonian frog Phyllomedusa bicolor. FEBS Lett. 302 (2), 151–154. doi:10.1016/0014-5793(92)80427-I
PubMed Abstract | CrossRef Full Text | Google Scholar
Moen, D. S., and Wiens, J. J. (2009). Phylogenetic evidence for competitively driven divergence: Body-size evolution in caribbean treefrogs (hylidae: Osteopilus). Evolution 63, 195–214. doi:10.1111/j.1558-5646.2008.00538.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Montecucchi, P. C., Henschen, A., and Erspamer, V. (1979). Structure of sauvagine, a vasoactive peptide from the skin of a frog. Struct. Activity Nat. Peptides 360, 1178. doi:10.1515/9783110861631
CrossRef Full Text | Google Scholar
Montecucchi, P. C., de Castiglione, R., Piani, S., Gozzini, L., and Erspamer, V. (1981a). Amino acid composition and sequence of dermorphin, a novel opiate‐like peptide from the skin of Phyllomedusa sauvagei. Int. J. Peptide Protein Res. 17 (3), 275–283. doi:10.1111/j.1399-3011.1981.tb01993.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Montecucchi, P. C., de Castiglione, R., and Erspamer, V. (1981b). Identification of dermorphin and Hyp6‐dermorphin in skin extracts of the Brazilian frog Phyllomedusa rhodei. Int. J. Peptide Protein Res. 17 (3), 316–321. doi:10.1111/j.1399-3011.1981.tb01997.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Mor, A., Van Huong, N., Delfour, A., Migliore-Samour, D., and Nicolas, P. (1991). Isolation, amino acid sequence and synthesis of dermaseptin, a novel antimicrobial peptide of amphibian skin. Biochemistry 30 (36), 8824–8830.
PubMed Abstract | CrossRef Full Text | Google Scholar
Mor, A., Amiche, M., and Nicolas, P. (1994a). Structure, synthesis, and activity of dermaseptin b, a novel vertebrate defensive peptide from frog skin: Relationship with adenoregulin. Biochemistry 33 (21), 6642–6650. doi:10.1021/bi00187a034
PubMed Abstract | CrossRef Full Text | Google Scholar
Mor, A., Chartrel, N., Vaudry, H., and Nicolas, P. (1994b). Skin peptide tyrosine-tyrosine, a member of the pancreatic polypeptide family: Isolation, structure, synthesis, and endocrine activity. Proc. Natl. Acad. Sci. 91 (22), 10295–10299. doi:10.1073/pnas.91.22.10295
PubMed Abstract | CrossRef Full Text | Google Scholar
Mor, A., Khaled, H., and Pierre, N. (1994c). The vertebrate peptide antibiotics dermaseptins have overlapping structural features but target specific microorganisms. J. Biol. Chem. 269 (50), 31635–31641. doi:10.1016/S0021-9258(18)31742-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Morais, D. R., Lanaro, R., Barbosa, I. L., Santos, J. M., Cunha, K. F., Hernandes, V. V., et al. (2018). Ayahuasca and Kambo intoxication after alternative natural therapy for depression, confirmed by mass spectrometry. Forensic Toxicol. 36 (1), 212–221. doi:10.1007/s11419-017-0394-5
CrossRef Full Text | Google Scholar
Mota, E. P., Kaefer, I. L., da Silva Nunes, M., Lima, A. P., and Farias, I. P. (2020). Hidden diversity within the broadly distributed Amazonian giant monkey frog (Phyllomedusa bicolor: Phyllomedusidae). Amphibia-Reptilia 41 (3), 349–359. doi:10.1163/15685381-bja10003
CrossRef Full Text | Google Scholar
Nakata, N., Sakurada, S., Sakurada, T., Kisara, K., Sasaki, Y., and Suzuki, K. (1986). Physical dependence of a dermorphin tetrapeptide analog, [D-Arg2, Sar4]-dermorphin (1–4) in the rat. Pharmacol. Biochem. Behav. 24 (1), 27–31. doi:10.1016/0091-3057(86)90039-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Naot, D., Musson, D. S., and Cornish, J. (2018). The activity of peptides of the calcitonin family in bone. Physiol. Rev. 99 (1), 781–805. doi:10.1152/physrev.00066.2017
PubMed Abstract | CrossRef Full Text | Google Scholar
Neckel-Oliveira, S., and Wachlevski, M. (2004). Predation on the arboreal eggs of three species of Phyllomedusa in Central Amazônia. Journal of Herpetology 38 (2), 244–248. doi:10.1670/162-03A
CrossRef Full Text | Google Scholar
Neckel-Sliveira, S., and Wachlevski, M. (2004). Predation on the arboreal eggs of three species of Phyllomedusa in Central Amazonia. J. Herpetology 38 (2), 244–248. doi:10.1670/162-03A
CrossRef Full Text | Google Scholar
Negri, L., Erspamer, G. F., Severini, C., Potenza, R. L., Melchiorri, P., and Erspamer, V. (1992). Dermorphin-related peptides from the skin of Phyllomedusa bicolor and their amidated analogs activate two μ opioid receptor subtypes that modulate antinociception and catalepsy in the rat. Proc. Natl. Acad. Sci. 89 (15), 7203–7207. doi:10.1073/pnas.89.15.7203
PubMed Abstract | CrossRef Full Text | Google Scholar
Nicolas, P., and Amiche, M. (2006). “The dermaseptins,” in Handbook of biologically active peptides (Academic Press), 295–304. doi:10.1016/B978-012369442-3/50048-9
CrossRef Full Text | Google Scholar
Nicolas, P., and El Amri, C. (2009). The dermaseptin superfamily: A genebased combinatorial library of antimicrobial peptides. Biochimica Biophysica Acta 1788 (8), 1537–1550. doi:10.1016/j.bbamem.2008.09.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Nicolas, P., and Mor, A. (1995). Peptides as weapons against microorganisms in the chemical defense system of vertebrates. Annu. Rev. Microbiol. 49, 277–304. doi:10.1146/annurev.mi.49.100195.001425
PubMed Abstract | CrossRef Full Text | Google Scholar
Ouboter, P. E., and Jairam, R. (2012). Amphibians of Suriname. Leiden, Boston: Brill.
Peleg Hasson, S., Shachar, E., Kessner, R., Shpigel, S., and Wolf, I. (2021). Kambô-induced systemic inflammatory response: A case report of acute disease progression of cholangiocarcinoma. Integr. Cancer Ther. 20, 1534735421999106. doi:10.1177/1534735421999106
CrossRef Full Text | Google Scholar
Pescatore, R., Marrone, G. F., Sedberry, S., Vinton, D., Finkelstein, N., Katlowitz, Y. E., et al. (2015). Synthesis and pharmacology of halogenated δ -opioid-selective [d-Ala(2)]deltorphin II peptide analogues. ACS Chem. Neurosci. 6 (6), 905–910. doi:10.1021/acschemneuro.5b00060
PubMed Abstract | CrossRef Full Text | Google Scholar
Pierre, T. N., Seon, A. A., Amiche, M., and Nicolas, P. (2000). Phylloxin, a novel peptide antibiotic of the dermaseptin family of antimicrobial/opioid peptide precursors. Eur. J. Biochem. 267 (2), 370–378. doi:10.1046/j.1432-1327.2000.01012.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Pinto, P., and Amésquita, A. (2022). Coloration, toxicity, and vertical distribution in larval phyllomedusine frogs: Is there an anti-predator syndrome? Authorea. doi:10.22541/au.164864450.06373836
CrossRef Full Text | Google Scholar
Pinto, R. M. C., Py-Daniel, S. S., and Menin, M. (2013). Redescription of the tadpole of Phyllomedusa bicolor (Anura: Hylidae) from central amazonia. S. Am. J. Herpetology 8 (1), 67–72. doi:10.2994/SAJH-D-13-00003.1
CrossRef Full Text | Google Scholar
Pogorzelska, J., and Łapiński, T. W. (2017). Toxic hepatitis caused by the excretions of the Phyllomedusa bicolor frog–a case report. Clin. Exp. Hepatology 3 (1), 33–34. doi:10.5114/ceh.2017.65228
PubMed Abstract | CrossRef Full Text | Google Scholar
Raaymakers, C., Verbrugghe, E., Stijlemans, B., Martel, A., Pasmans, F., and Roelants, K. (2018). The anuran skin peptide bradykinin mediates its own absorption across epithelial barriers of the digestive tract. Peptides 103, 84–89. doi:10.1016/j.peptides.2018.03.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Resende, J. M., Moraes, C. M., Prates, M. V., Cesar, A., Almeida, F. C., Mundim, N. C., et al. (2008). Solution NMR structures of the antimicrobial peptides phylloseptin-1, -2, and-3 and biological activity: The role of charges and hydrogen bonding interactions in stabilizing helix conformations. Peptides 29 (10), 1633–1644. doi:10.1016/j.peptides.2008.06.022
PubMed Abstract | CrossRef Full Text | Google Scholar
Ribeiro, F. S. (2017). O Kambô na rede: Divulgação de uma prática tradicional indígena na internet [Monography]. [Curitiba (PR)]. Federal University of Paraná.
Richter, K., Egger, R., and Kreil, G. (1990). Molecular cloning of a cDNA encoding the bombesin precursor in skin of Bombina variegata. FEBS Lett. 262 (2), 353–355. doi:10.1016/0014-5793(90)80227-A
PubMed Abstract | CrossRef Full Text | Google Scholar
Rocha, S. M. C. da, Lima, A. P., and Kaefer, I. L. (2021). Key roles of paternal care and climate on offspring survival of an Amazonian poison frog. An. Acad. Bras. Ciências 93 (3). doi:10.1590/0001-3765202120210067
CrossRef Full Text | Google Scholar
Rödel, M. O., Brede, C., Hirschfeld, M., Schmitt, T., Favreau, P., Stöcklin, R., et al. (2013). Chemical camouflage–a frog’s strategy to co-exist with aggressive ants. PLoS One 8 (12), e81950. doi:10.1371/journal.pone.0081950
PubMed Abstract | CrossRef Full Text | Google Scholar
Rodríguez, A., Poth, D., Schulz, S., and Vences, M. (2011). Discovery of skin alkaloids in a miniaturized eleutherodactylid frog from Cuba. Biol. Lett. 7 (3), 414–418. doi:10.1098/rsbl.2010.0844
PubMed Abstract | CrossRef Full Text | Google Scholar
Rodríguez, L. O., and Duellman, W. E. (1994). Guide to the frogs of the iquitos region, amazonian Peru: Lawrence: Associacion de ecologia y conservacion, Amazon Center for environmental education and research. Natural History Museum, University of Kansas.
Roy, R., Baranwal, A., and Espiridion, E. D. (2018). Can overuse of kambô cause psychosis? Cureus 10 (6), e2770. doi:10.7759/cureus.2770
PubMed Abstract | CrossRef Full Text | Google Scholar
Rudgley, R. (1993). Essential substances: A cultural history of intoxicants in society, 195.
Ruibal, R., and Shoemaker, V. (1984). Osteoderms in anurans. J. Herpetology 18 (3), 313–328. doi:10.2307/1564085
CrossRef Full Text | Google Scholar
Sacco, M. A., Zibetti, A., Bonetta, C. F., Scalise, C., Abenavoli, L., Guarna, F., et al. (2022). Kambo: Natural drug or potential toxic agent? A literature review of acute poisoning cases. Toxicol. Rep. 9, 905–913. doi:10.1016/j.toxrep.2022.04.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Santos, C., Hamadat, S., Le Saux, K., Newton, C., Mazouni, M., Zargarian, L., et al. (2017). Studies of the antitumor mechanism of action of dermaseptin B2, a multifunctional cationic antimicrobial peptide, reveal a partial implication of cell surface glycosaminoglycans. PLoS One 12 (8), e0182926. doi:10.1371/journal.pone.0182926
PubMed Abstract | CrossRef Full Text | Google Scholar
Saporito, R. A., Donnelly, M. A., Spande, T. F., and Garraffo, H. M. (2012). A review of chemical ecology in poison frogs. Chemoecology 22 (3), 159–168. doi:10.1007/s00049-011-0088-0
CrossRef Full Text | Google Scholar
Saporito, R. A., Spande, T. F., and Garraffo, H. M. (2009). Arthropod alkaloids in poison frogs: A review of the ‘dietary hypothesis. Heterocycles 79 (1), 277–297. doi:10.3987/REV-08-SR(D)11
CrossRef Full Text | Google Scholar
Sazima, I. (1974). Experimental predation on the leaf-frog Phyllomedusa rohdei by the water snake Liophis militaris. J. Herpetology 8 (4), 376–377.
CrossRef Full Text | Google Scholar
Schäffer, D. A., and Gábriel, R. (2005). Two major tachykinins, substance P and substance K, are localized to distinct subsets of amacrine cells in the anuran retina. Neurosci. Lett. 386 (3), 194–198. doi:10.1016/j.neulet.2005.06.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Schmidt, T. T., Reiche, S., Hage, C. L., Bermpohl, F., and Majić, T. (2020). Acute and subacute psychoactive effects of kambô, the secretion of the amazonian giant maki frog (Phyllomedusa bicolor): Retrospective reports. Sci. Rep. 10 (1), 1–11. doi:10.1038/s41598-020-78527-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Seon, A. A., Pierre, T. N., Redeker, V., Lacombe, C., Delfour, A., Nicolas, P., et al. (2000). Isolation, structure, synthesis, and activity of a new member of the calcitonin gene-related peptide family from frog skin and molecular cloning of its precursor. J. Biol. Chem. 275 (8), 5934–5940. doi:10.1074/jbc.275.8.5934
PubMed Abstract | CrossRef Full Text | Google Scholar
Severini, C., Improta, G., Falconieri-Erspamer, G., Salvadori, S., and Erspamer, V. (2002). The tachykinin peptide family. Pharmacol. Rev. 54 (2), 285–322. doi:10.1124/pr.54.2.285
PubMed Abstract | CrossRef Full Text | Google Scholar
Shai, Y. (1995). Molecular recognition between membrane-spanning polypeptides. Trends Biochem. Sci. 20 (11), 460–464. doi:10.1016/S0968-0004(00)89101-X
PubMed Abstract | CrossRef Full Text | Google Scholar
Shine, R. (1979). Sexual selection and sexual dimorphism in Amphibia. Copeia 1979 (2), 297–306. doi:10.2307/1443418
CrossRef Full Text | Google Scholar
Shiva, V. (1991). Biotechnology development and conservation of biodiversity. Econ. Political Wkly. 26 (48), 2740–2746.
Silva, F. V. A. D., Monteiro, W. M., and Bernarde, P. S. (2019). Kambô” frog (Phyllomedusa bicolor): Use in folk medicine and potential health risks. Rev. Soc. Bras. Med. Trop. 52. doi:10.1590/0037-8682-0467-2018
PubMed Abstract | CrossRef Full Text | Google Scholar
Silva, Y. B. S., Almeida-Santos, W., Araújo, A. S., and Costa-Campos, C. E. (2020). Notes on the reproductive biology of Phyllomedusa bicolor (Anura, Phyllomedusidae) in the Amazon forest of northern Brazil. Herpetol. Notes 13, 931–935.
Souza, M. B. (2009). Anfíbios: Reserva Extrativista do Alto Juruá e Parque Nacional da Serra do Divisor, Acre. Campinas: Federal University of Acre, 77.
Souza, M. B., Aquino, T. V., Lima, E. C., and Mendes, M. K. (2002). “Anfíbios,” in Enciclopédia da floresta: O alto Juruá: Práticas e conhecimento das populações. 1ed. Editors M. C. Cunha, and M. B. Almeida Orgs (São Paulo: Companhia das Letras), 601-614.
Steinhoff, M. S., von Mentzer, B., Geppetti, P., Pothoulakis, C., and Bunnett, N. W. (2014). Tachykinins and their receptors: Contributions to physiological control and the mechanisms of disease. Physiol. Rev. 94 (1), 265–301. doi:10.1152/physrev.00031.2013
PubMed Abstract | CrossRef Full Text | Google Scholar
Tastevin, C. (1925). Le fleuve murú: Ses habitants: Croyances et moeurs kachinaua. Société Géogr. 43, 401–422.
Thibaudeau, G., and Altig, R. (2012). Coloration of anuran tadpoles (Amphibia): Development, dynamics, function, and hypotheses. Int. Sch. Res. Netw. Zoology, 1–16. doi:10.5402/2012/725203
CrossRef Full Text | Google Scholar
Thompson, C., Malcolm, B., and Tegzes, J. (2022). Use of Phyllomedusa bicolour secretion during kambô ritual: Observational responses, dosage, and risk of adverse events. Toxicol. Commun. 6 (1), 6–12. doi:10.1177/23978473221085746
CrossRef Full Text | Google Scholar
Thompson, C., and Williams, M. L. (2022). Review of the physiological effects of Phyllomedusa bicolor skin secretion peptides on humans receiving Kambô. Toxicol. Res. Appl. 6, 23978473221085746.
CrossRef Full Text | Google Scholar
Tiwari, V. (2019). Post-translational modification of ESKAPE pathogens as a potential target in drug discovery. Drug Discov. Today 24 (3), 814–822. doi:10.1016/j.drudis.2018.12.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Toledo, L. F., Sazima, I., and Haddad, C. F. (2011). Behavioural defences of anurans: An overview. Ethol. Ecol. Evol. 23 (1), 1–25. doi:10.1080/03949370.2010.534321
CrossRef Full Text | Google Scholar
Toledo, R. C., and Jared, C. (1993). Cutaneous adaptations to water balance in amphibians. Comp. Biochem. Physiology Part A Physiology 105 (4), 593–608. doi:10.1016/0300-9629(93)90259-7
CrossRef Full Text | Google Scholar
Toledo, R. D., and Jared, C. (1995). Cutaneous granular glands and amphibian venoms. Comp. Biochem. Physiology Part A Physiology 111 (1), 1–29. doi:10.1016/0300-9629(95)98515-I
CrossRef Full Text | Google Scholar
Tsuji-Nishikido, B. M., and Menin, M. (2011). Distribution of frogs in riparian areas of an urban forest fragment in Central Amazonia. Biota Neotropica 11 (2), 63–70. doi:10.1590/S1676-06032011000200007
CrossRef Full Text | Google Scholar
van Zoggel, H., Hamma-Kourbali, Y., Galanth, C., Ladram, A., Nicolas, P., Courty, J., et al. (2012). Antitumor and angiostatic peptides from frog skin secretions. Amino Acids 42 (1), 385–395. doi:10.1007/s00726-010-0815-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Vanhoye, D., Bruston, F., El Amri, S., Ladram, A., Amiche, M., and Nicolas, P. (2004). Membrane association, electrostatic sequestration, and cytotoxicity of Gly-Leu-rich peptide orthologs with differing functions. Biochemistry 43 (26), 8391–8409. doi:10.1021/bi0493158
PubMed Abstract | CrossRef Full Text | Google Scholar
Venâncio, N. M., and Melo-Sampaio, P. R. (2010). Reproductive behavior of the giant leaf frog Phyllomedusa bicolor (Anura: Hylidae) in the Western Amazon. Phyllomedusa J. Herpetology 9 (1), 63–67. doi:10.11606/issn.2316-9079.v9i1p63-67
CrossRef Full Text | Google Scholar
Wager, V. (1986). Frogs of South Africa: Their fascinating life stories. Johannesburg: Delta.
Wagler, J. (1830). Adaptative plasticity in hatching age: A response to predation risk tradeoffs Natürliches system der Amphibien, mit vorangehender Classification der Säugthiere und Vägel. München, J.G. Cotta vi + 354
Warkentin, K. M. (1995). Adaptative plasticity in hatching age: A response to predation risk tradeoffs. Proc. Natl. Acad. Sci. 92 (8), 3507–3510. doi:10.1073/pnas.92.8.3507
PubMed Abstract | CrossRef Full Text | Google Scholar
Wiens, J. J., Fetzner, J. W., Parkinson, C. L., and Reeder, T. W. (2005). Hylid frog phylogeny and sampling strategies for speciose clades. Syst. Biol. 54 (5), 719–748. doi:10.1080/10635150500234625
PubMed Abstract | CrossRef Full Text | Google Scholar
Wiens, J. J., Graham, C. H., Moen, D. S., Smith, S. A., and Reeder, T. W. (2006). Evolutionary and ecological causes of the latitudinal diversity gradient in hylid frogs: Treefrog trees unearth the roots of high tropical diversity. Am. Nat. 168, 579–596. doi:10.1086/507882
PubMed Abstract | CrossRef Full Text | Google Scholar
Williams, C. R., Brodie, E. D., Tyler, M. J., and Walker, S. J. (2000). Antipredator mechanisms of Australian frogs. J. Herpetology 34 (3), 431–443. doi:10.2307/1565367
CrossRef Full Text | Google Scholar
Zhang, R., Zhou, M., Wang, L., McGrath, S., Chen, T., Chen, X., et al. (2010). Phylloseptin-1 (PSN-1) from Phyllomedusa sauvagei skin secretion: A novel broad-spectrum antimicrobial peptide with antibiofilm activity. Mol. Immunol. 47 (11-12), 2030–2037. doi:10.1016/j.molimm.2010.04.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Оригинал текста: https://www.frontiersin.org/articles/10.3389/fphar.2022.997318/full
